

Adaptive NK Cells Resist Regulatory T-cell Suppression Driven by IL37
- PMID: 29784636
- PMCID: PMC6030483
- DOI: 10.1158/2326-6066.CIR-17-0498
Adaptive NK Cells Resist Regulatory T-cell Suppression Driven by IL37
Abstract
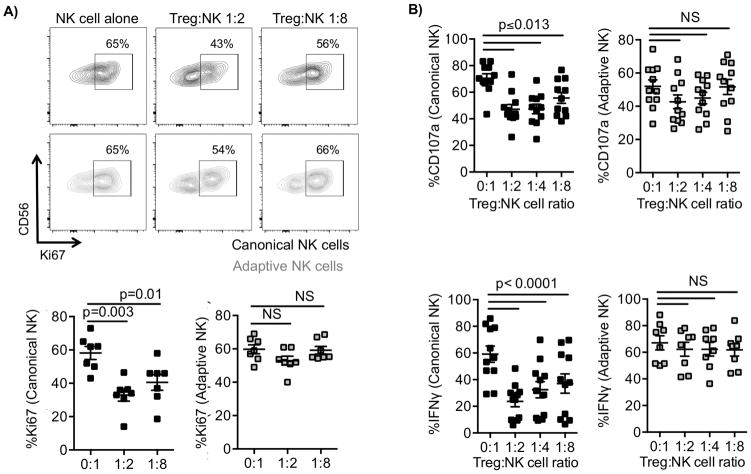
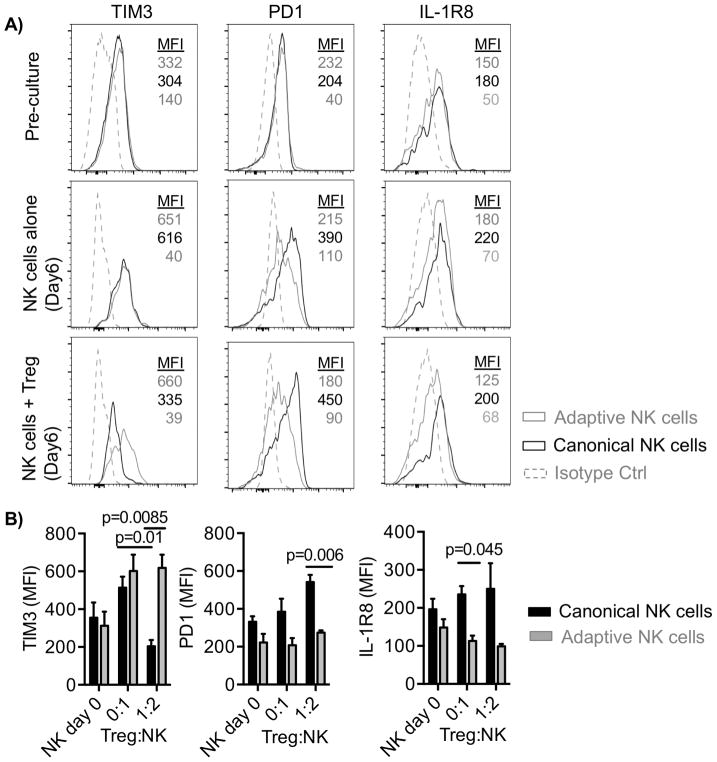
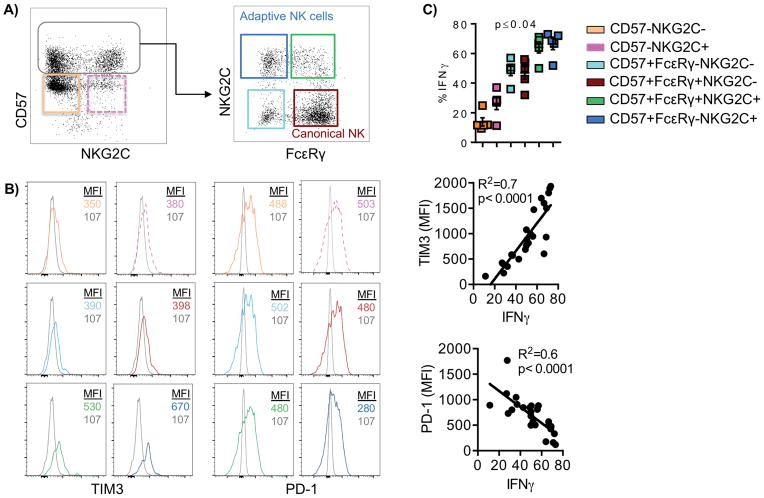
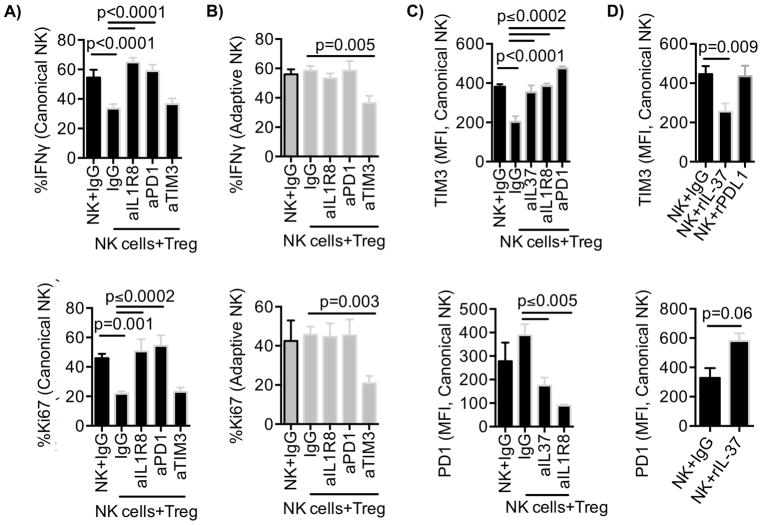
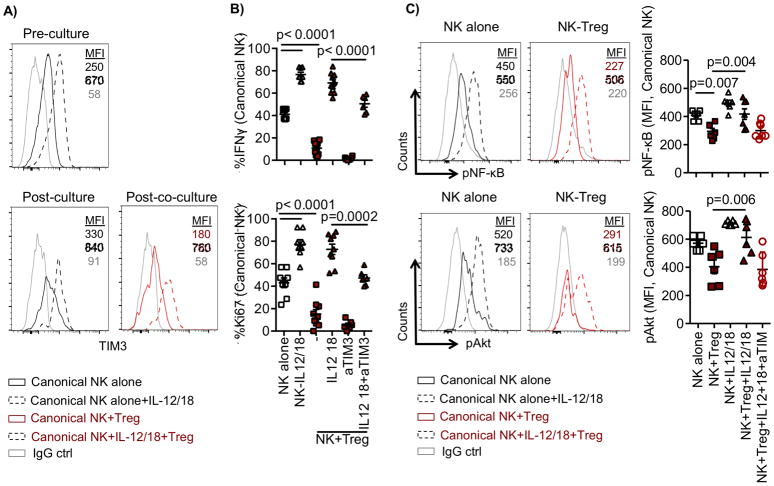
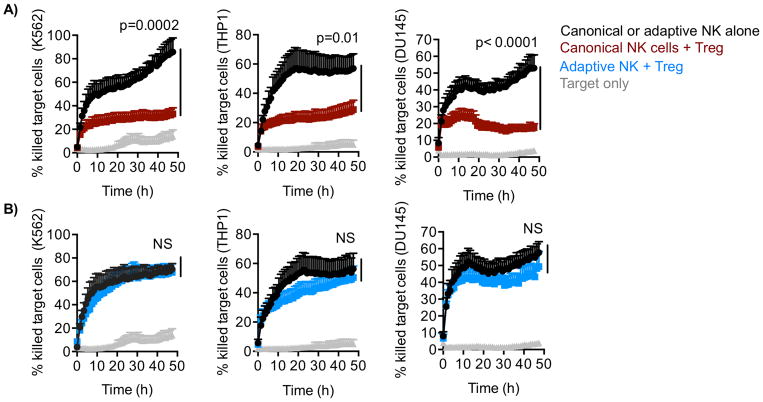
Natural killer (NK) cells are capable of fighting viral infections and cancer. However, these responses are inhibited by immune suppressor cells in the tumor microenvironment. Tumor progression promotes the recruitment and generation of intratumoral regulatory T cells (Treg), associated with a poor prognosis in cancer patients. Here, we show that canonical NK cells are highly susceptible to Treg-mediated suppression, in contrast to highly resistant CD57+ FcεRγ-NKG2C+ adaptive (CD56+CD3-) NK cells that expand in cytomegalovirus exposed individuals. Specifically, Tregs suppressed canonical but not adaptive NK-cell proliferation, IFNγ production, degranulation, and cytotoxicity. Treg-mediated suppression was associated with canonical NK-cell downregulation of TIM3, a receptor that activates NK-cell IFNγ production upon ligand engagement, and upregulation of the NK-cell inhibitory receptors PD-1 and the IL1 receptor family member, IL1R8 (SIGIRR or TIR8). Treg production of the IL1R8 ligand, IL37, contributed to the phenotypic changes and diminished function in Treg-suppressed canonical NK cells. Blocking PD-1, IL1R8, or IL37 abrogated Treg suppression of canonical NK cells while maintaining NK-cell TIM3 expression. Our data uncover new mechanisms of Treg-mediated suppression of canonical NK cells and identify that adaptive NK cells are inherently resistant to Treg suppression. Strategies to enhance the frequency of adaptive NK cells in the tumor microenvironment or to blunt Treg suppression of canonical NK cells will enhance the efficacy of NK-cell cancer immunotherapy. Cancer Immunol Res; 6(7); 766-75. ©2018 AACR.
©2018 American Association for Cancer Research.
Conflict of interest statement
Figures
Similar articles
-
Surface Immune Checkpoints as Potential Biomarkers in Physiological Pregnancy and Recurrent Pregnancy Loss.Int J Mol Sci. 2024 Aug 29;25(17):9378. doi: 10.3390/ijms25179378. Int J Mol Sci. 2024. PMID: 39273326 Free PMC article.
-
Late Development of FcεRγneg Adaptive Natural Killer Cells Upon Human Cytomegalovirus Reactivation in Umbilical Cord Blood Transplantation Recipients.Front Immunol. 2018 May 15;9:1050. doi: 10.3389/fimmu.2018.01050. eCollection 2018. Front Immunol. 2018. PMID: 29868012 Free PMC article.
-
IL2 Variant Circumvents ICOS+ Regulatory T-cell Expansion and Promotes NK Cell Activation.Cancer Immunol Res. 2016 Nov;4(11):983-994. doi: 10.1158/2326-6066.CIR-15-0195. Epub 2016 Oct 3. Cancer Immunol Res. 2016. PMID: 27697858
-
PD/1-PD-Ls Checkpoint: Insight on the Potential Role of NK Cells.Front Immunol. 2019 Jun 4;10:1242. doi: 10.3389/fimmu.2019.01242. eCollection 2019. Front Immunol. 2019. PMID: 31214193 Free PMC article. Review.
-
The role of regulatory T cells in the control of natural killer cells: relevance during tumor progression.Immunol Rev. 2006 Dec;214:229-38. doi: 10.1111/j.1600-065X.2006.00445.x. Immunol Rev. 2006. PMID: 17100888 Review.
Cited by
-
Role of Interleukin-1 family in bone metastasis of prostate cancer.Front Oncol. 2022 Sep 27;12:951167. doi: 10.3389/fonc.2022.951167. eCollection 2022. Front Oncol. 2022. PMID: 36237303 Free PMC article. Review.
-
Natural Killer Cells Are Key Host Immune Effector Cells Affecting Survival in Autologous Peripheral Blood Hematopoietic Stem Cell Transplantation.Cells. 2022 Nov 2;11(21):3469. doi: 10.3390/cells11213469. Cells. 2022. PMID: 36359863 Free PMC article. Review.
-
NK cells are never alone: crosstalk and communication in tumour microenvironments.Mol Cancer. 2023 Feb 16;22(1):34. doi: 10.1186/s12943-023-01737-7. Mol Cancer. 2023. PMID: 36797782 Free PMC article. Review.
-
Phenotypic and Functional Analysis of Human NK Cell Subpopulations According to the Expression of FcεRIγ and NKG2C.Front Immunol. 2019 Dec 6;10:2865. doi: 10.3389/fimmu.2019.02865. eCollection 2019. Front Immunol. 2019. PMID: 31867015 Free PMC article.
-
Optimizing NK Cell-Based Immunotherapy in Myeloid Leukemia: Abrogating an Immunosuppressive Microenvironment.Front Immunol. 2021 Jun 17;12:683381. doi: 10.3389/fimmu.2021.683381. eCollection 2021. Front Immunol. 2021. PMID: 34220833 Free PMC article. Review.
References
-
- Liotta F, Gacci M, Frosali F, Querci V, Vittori G, Lapini A, et al. Frequency of regulatory T cells in peripheral blood and in tumour-infiltrating lymphocytes correlates with poor prognosis in renal cell carcinoma. BJU international. 2011;107(9):1500–6. doi: 10.1111/j.1464-410X.2010.09555.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R00 HL123638/HL/NHLBI NIH HHS/United States
- R01 HL118979/HL/NHLBI NIH HHS/United States
- R37 AI034495/AI/NIAID NIH HHS/United States
- R01 HL122216/HL/NHLBI NIH HHS/United States
- R35 CA197292/CA/NCI NIH HHS/United States
- R01 CA072669/CA/NCI NIH HHS/United States
- K99 HL123638/HL/NHLBI NIH HHS/United States
- UL1 TR002494/TR/NCATS NIH HHS/United States
- P01 CA065493/CA/NCI NIH HHS/United States
- R37 HL056067/HL/NHLBI NIH HHS/United States
- R01 HL056067/HL/NHLBI NIH HHS/United States
- P01 CA111412/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
