

Structure and Function of the 26S Proteasome
- PMID: 29652515
- PMCID: PMC6422034
- DOI: 10.1146/annurev-biochem-062917-011931
Structure and Function of the 26S Proteasome
Abstract
As the endpoint for the ubiquitin-proteasome system, the 26S proteasome is the principal proteolytic machine responsible for regulated protein degradation in eukaryotic cells. The proteasome's cellular functions range from general protein homeostasis and stress response to the control of vital processes such as cell division and signal transduction. To reliably process all the proteins presented to it in the complex cellular environment, the proteasome must combine high promiscuity with exceptional substrate selectivity. Recent structural and biochemical studies have shed new light on the many steps involved in proteasomal substrate processing, including recognition, deubiquitination, and ATP-driven translocation and unfolding. In addition, these studies revealed a complex conformational landscape that ensures proper substrate selection before the proteasome commits to processive degradation. These advances in our understanding of the proteasome's intricate machinery set the stage for future studies on how the proteasome functions as a major regulator of the eukaryotic proteome.
Keywords: 26S proteasome; AAA+ ATPase; deubiquitination; energy-dependent protein degradation; ubiquitin code; ubiquitin receptor.
Figures
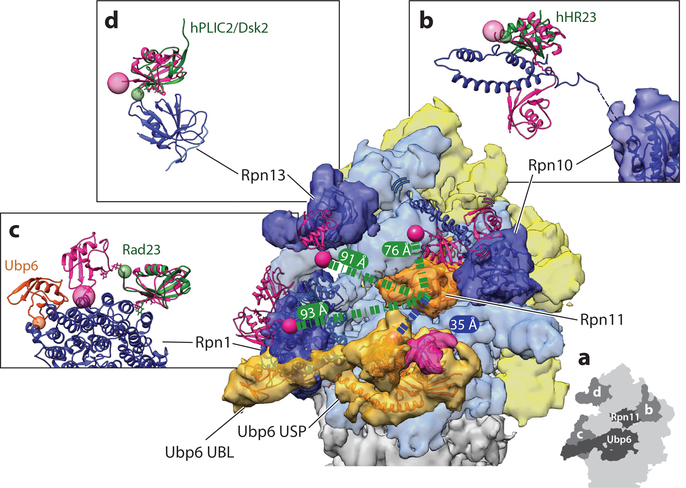
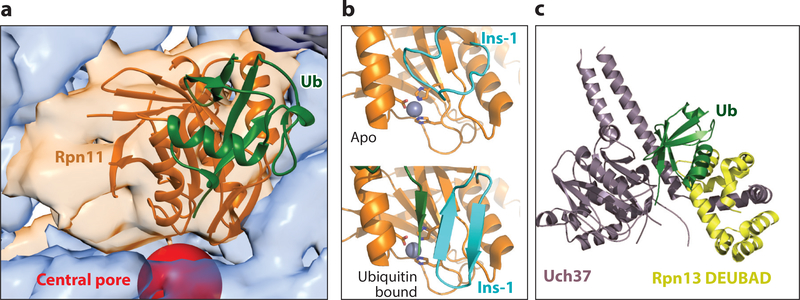
Similar articles
-
Specific lid-base contacts in the 26s proteasome control the conformational switching required for substrate degradation.Elife. 2019 Nov 28;8:e49806. doi: 10.7554/eLife.49806. Elife. 2019. PMID: 31778111 Free PMC article.
-
An AAA Motor-Driven Mechanical Switch in Rpn11 Controls Deubiquitination at the 26S Proteasome.Mol Cell. 2017 Sep 7;67(5):799-811.e8. doi: 10.1016/j.molcel.2017.07.023. Epub 2017 Aug 24. Mol Cell. 2017. PMID: 28844860
-
An empirical energy landscape reveals mechanism of proteasome in polypeptide translocation.Elife. 2022 Jan 20;11:e71911. doi: 10.7554/eLife.71911. Elife. 2022. PMID: 35050852 Free PMC article.
-
Structure, Dynamics and Function of the 26S Proteasome.Subcell Biochem. 2021;96:1-151. doi: 10.1007/978-3-030-58971-4_1. Subcell Biochem. 2021. PMID: 33252727 Review.
-
Deubiquitination Reactions on the Proteasome for Proteasome Versatility.Int J Mol Sci. 2020 Jul 27;21(15):5312. doi: 10.3390/ijms21155312. Int J Mol Sci. 2020. PMID: 32726943 Free PMC article. Review.
Cited by
-
How Do Post-Translational Modifications Influence the Pathomechanistic Landscape of Huntington's Disease? A Comprehensive Review.Int J Mol Sci. 2020 Jun 16;21(12):4282. doi: 10.3390/ijms21124282. Int J Mol Sci. 2020. PMID: 32560122 Free PMC article. Review.
-
Hiding the Elephant in the Room with Experimental Neutrons.Biophys J. 2020 Jul 21;119(2):234-235. doi: 10.1016/j.bpj.2020.05.038. Epub 2020 Jun 24. Biophys J. 2020. PMID: 32640187 Free PMC article. No abstract available.
-
Specific lid-base contacts in the 26s proteasome control the conformational switching required for substrate degradation.Elife. 2019 Nov 28;8:e49806. doi: 10.7554/eLife.49806. Elife. 2019. PMID: 31778111 Free PMC article.
-
Proteasome serves as pivotal regulator in Angiostrongylus cantonensis-induced eosinophilic meningoencephalitis.PLoS One. 2019 Aug 15;14(8):e0220503. doi: 10.1371/journal.pone.0220503. eCollection 2019. PLoS One. 2019. PMID: 31415587 Free PMC article.
-
HUWE1 controls tristetraprolin proteasomal degradation by regulating its phosphorylation.Elife. 2023 Mar 24;12:e83159. doi: 10.7554/eLife.83159. Elife. 2023. PMID: 36961408 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
