

SLAM-seq defines direct gene-regulatory functions of the BRD4-MYC axis
- PMID: 29622725
- PMCID: PMC6409205
- DOI: 10.1126/science.aao2793
SLAM-seq defines direct gene-regulatory functions of the BRD4-MYC axis
Abstract
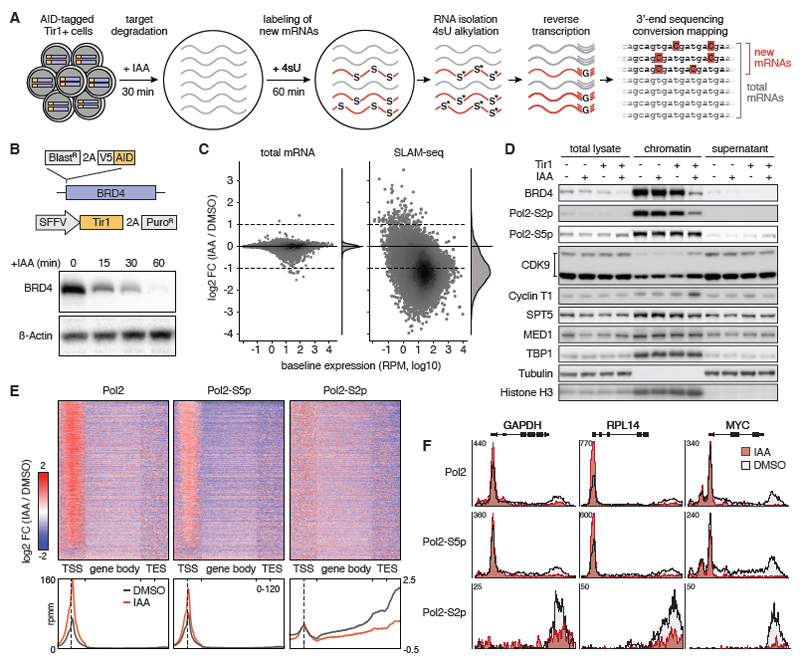
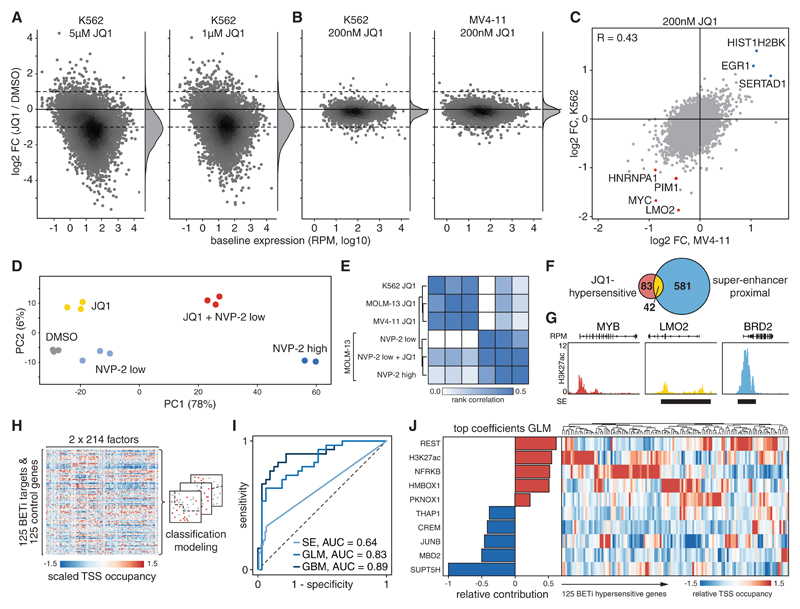
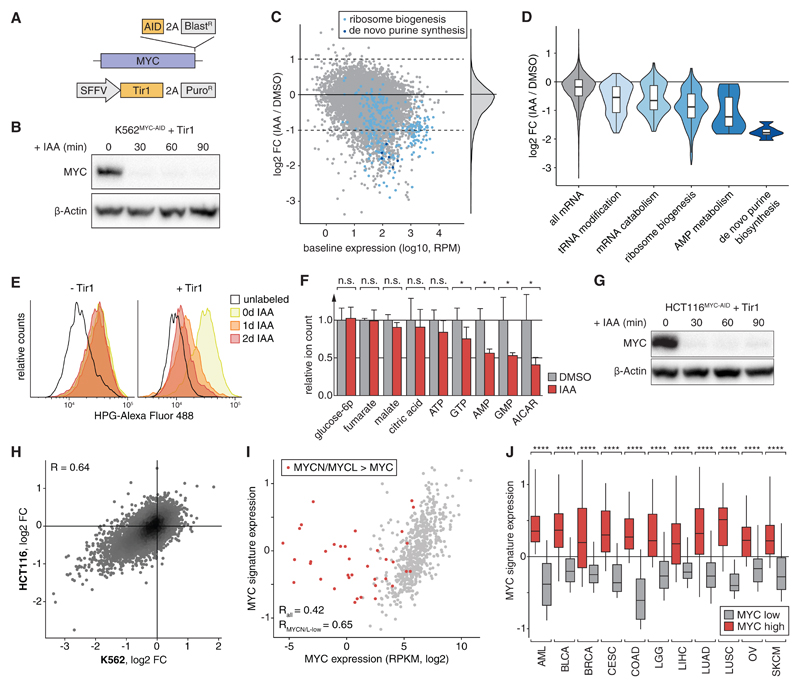
Defining direct targets of transcription factors and regulatory pathways is key to understanding their roles in physiology and disease. We combined SLAM-seq [thiol(SH)-linked alkylation for the metabolic sequencing of RNA], a method for direct quantification of newly synthesized messenger RNAs (mRNAs), with pharmacological and chemical-genetic perturbation in order to define regulatory functions of two transcriptional hubs in cancer, BRD4 and MYC, and to interrogate direct responses to BET bromodomain inhibitors (BETis). We found that BRD4 acts as general coactivator of RNA polymerase II-dependent transcription, which is broadly repressed upon high-dose BETi treatment. At doses triggering selective effects in leukemia, BETis deregulate a small set of hypersensitive targets including MYC. In contrast to BRD4, MYC primarily acts as a selective transcriptional activator controlling metabolic processes such as ribosome biogenesis and de novo purine synthesis. Our study establishes a simple and scalable strategy to identify direct transcriptional targets of any gene or pathway.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Taking aim at transcriptional regulator targets.Nat Rev Genet. 2018 Jun;19(6):328. doi: 10.1038/s41576-018-0010-5. Nat Rev Genet. 2018. PMID: 29686398 No abstract available.
-
SLAMing transcription.Nat Chem Biol. 2018 Jun;14(6):525. doi: 10.1038/s41589-018-0074-8. Nat Chem Biol. 2018. PMID: 29769731 No abstract available.
-
BRD4 and MYC-clarifying regulatory specificity.Science. 2018 May 18;360(6390):713-714. doi: 10.1126/science.aat6664. Science. 2018. PMID: 29773735 No abstract available.
Similar articles
-
AZD5153: A Novel Bivalent BET Bromodomain Inhibitor Highly Active against Hematologic Malignancies.Mol Cancer Ther. 2016 Nov;15(11):2563-2574. doi: 10.1158/1535-7163.MCT-16-0141. Epub 2016 Aug 29. Mol Cancer Ther. 2016. PMID: 27573426
-
BRD4 and MYC: power couple in transcription and disease.FEBS J. 2023 Oct;290(20):4820-4842. doi: 10.1111/febs.16580. Epub 2022 Aug 3. FEBS J. 2023. PMID: 35866356 Free PMC article. Review.
-
Highly active combination of BRD4 antagonist and histone deacetylase inhibitor against human acute myelogenous leukemia cells.Mol Cancer Ther. 2014 May;13(5):1142-54. doi: 10.1158/1535-7163.MCT-13-0770. Epub 2014 Jan 16. Mol Cancer Ther. 2014. PMID: 24435446
-
Dissecting the impact of bromodomain inhibitors on the Interferon Regulatory Factor 4-MYC oncogenic axis in multiple myeloma.Hematol Oncol. 2022 Aug;40(3):417-429. doi: 10.1002/hon.3016. Epub 2022 May 18. Hematol Oncol. 2022. PMID: 35544413 Free PMC article.
-
MYC as a regulator of ribosome biogenesis and protein synthesis.Nat Rev Cancer. 2010 Apr;10(4):301-9. doi: 10.1038/nrc2819. Nat Rev Cancer. 2010. PMID: 20332779 Review.
Cited by
-
RNA kinetics influence the response to transcriptional perturbation in leukaemia cell lines.NAR Cancer. 2024 Oct 3;6(4):zcae039. doi: 10.1093/narcan/zcae039. eCollection 2024 Dec. NAR Cancer. 2024. PMID: 39372038 Free PMC article.
-
BRD4: a general regulator of transcription elongation.Transcription. 2022 Feb-Jun;13(1-3):70-81. doi: 10.1080/21541264.2022.2108302. Epub 2022 Sep 1. Transcription. 2022. PMID: 36047906 Free PMC article. Review.
-
BRD4 Prevents R-Loop Formation and Transcription-Replication Conflicts by Ensuring Efficient Transcription Elongation.Cell Rep. 2020 Sep 22;32(12):108166. doi: 10.1016/j.celrep.2020.108166. Cell Rep. 2020. PMID: 32966794 Free PMC article.
-
Nanoscale imaging of DNA-RNA identifies transcriptional plasticity at heterochromatin.Life Sci Alliance. 2024 Sep 17;7(12):e202402849. doi: 10.26508/lsa.202402849. Print 2024 Dec. Life Sci Alliance. 2024. PMID: 39288993 Free PMC article.
-
Nascent RNA analyses: tracking transcription and its regulation.Nat Rev Genet. 2019 Dec;20(12):705-723. doi: 10.1038/s41576-019-0159-6. Epub 2019 Aug 9. Nat Rev Genet. 2019. PMID: 31399713 Free PMC article. Review.
References
-
- Jones PA, Issa J-PJ, Baylin S. Targeting the cancer epigenome for therapy. Nat Rev Genet. 2016;17:630–641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
