

HSV-1-induced disruption of transcription termination resembles a cellular stress response but selectively increases chromatin accessibility downstream of genes
- PMID: 29579120
- PMCID: PMC5886697
- DOI: 10.1371/journal.ppat.1006954
HSV-1-induced disruption of transcription termination resembles a cellular stress response but selectively increases chromatin accessibility downstream of genes
Abstract
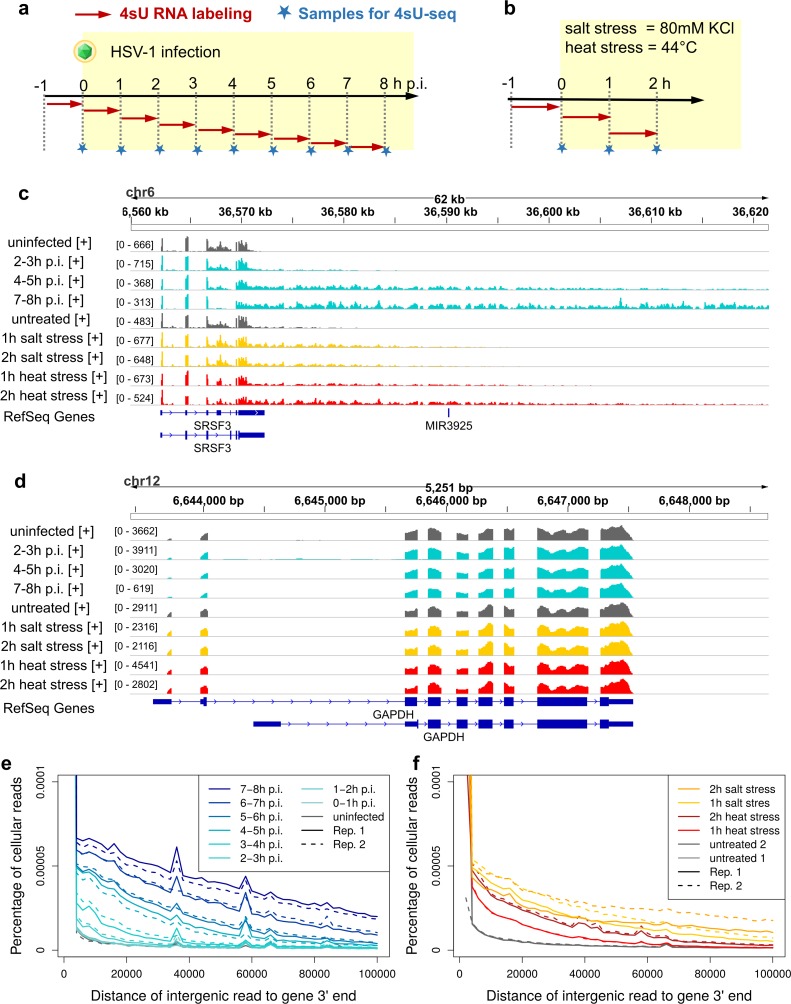
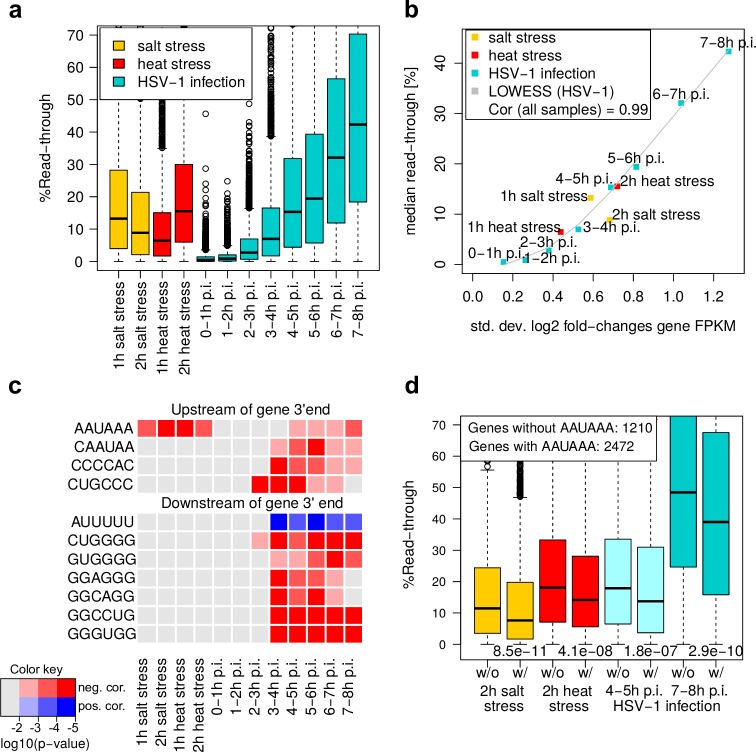
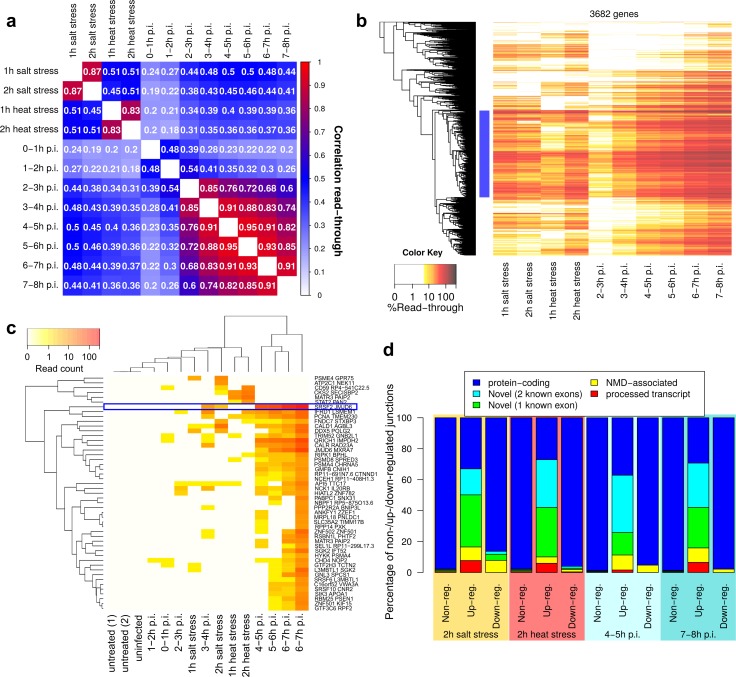
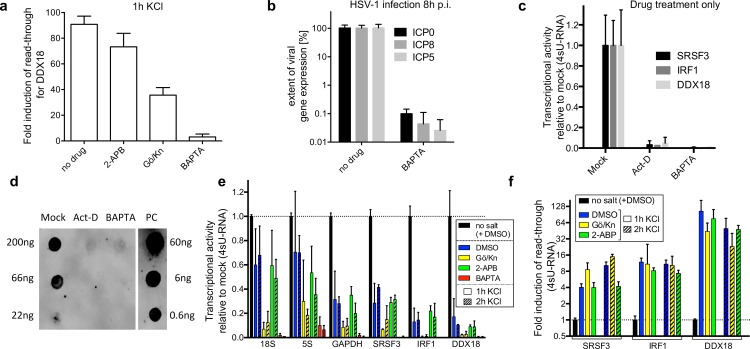
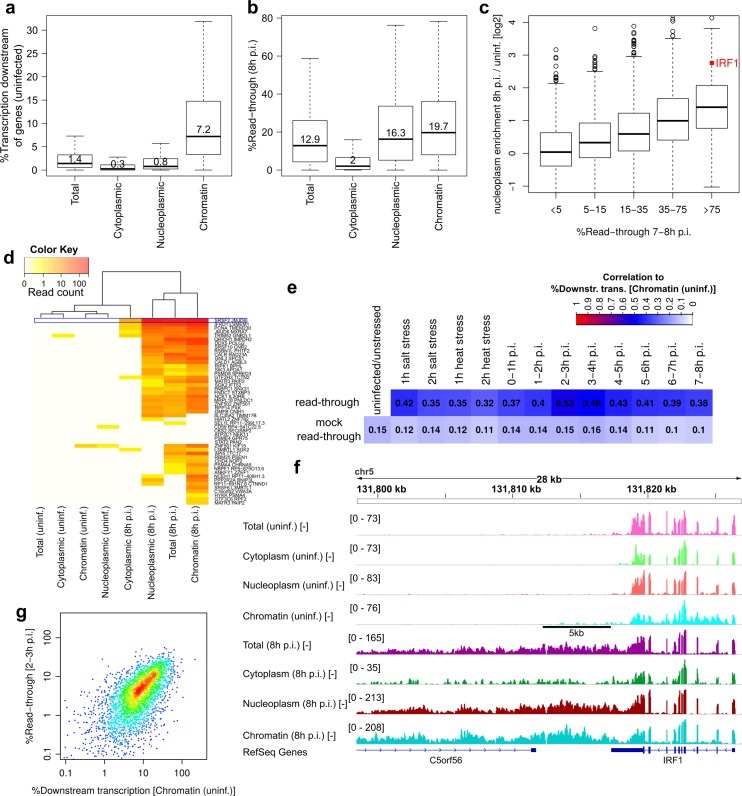
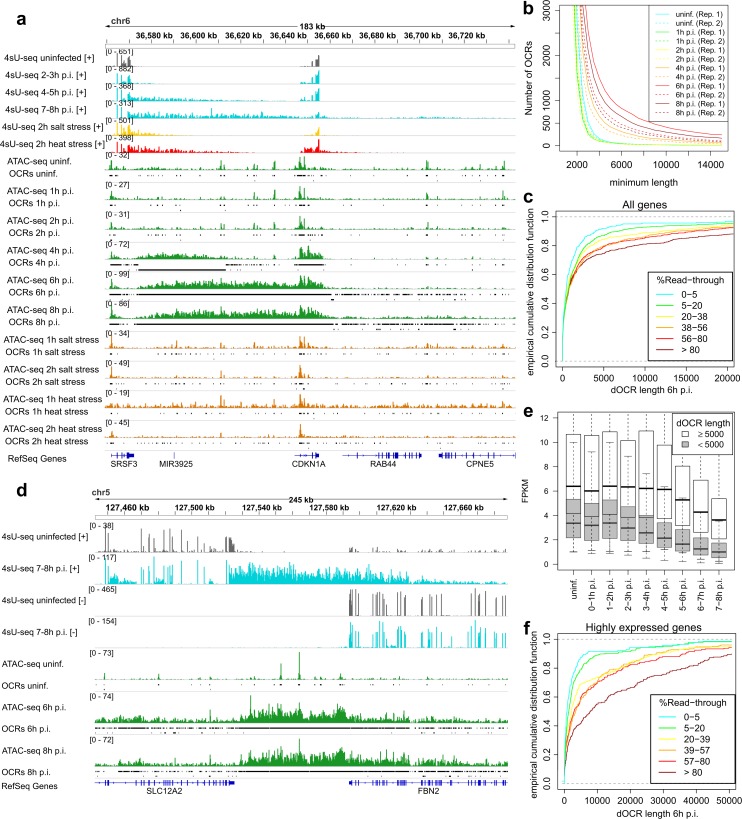
Lytic herpes simplex virus 1 (HSV-1) infection triggers disruption of transcription termination (DoTT) of most cellular genes, resulting in extensive intergenic transcription. Similarly, cellular stress responses lead to gene-specific transcription downstream of genes (DoG). In this study, we performed a detailed comparison of DoTT/DoG transcription between HSV-1 infection, salt and heat stress in primary human fibroblasts using 4sU-seq and ATAC-seq. Although DoTT at late times of HSV-1 infection was substantially more prominent than DoG transcription in salt and heat stress, poly(A) read-through due to DoTT/DoG transcription and affected genes were significantly correlated between all three conditions, in particular at earlier times of infection. We speculate that HSV-1 either directly usurps a cellular stress response or disrupts the transcription termination machinery in other ways but with similar consequences. In contrast to previous reports, we found that inhibition of Ca2+ signaling by BAPTA-AM did not specifically inhibit DoG transcription but globally impaired transcription. Most importantly, HSV-1-induced DoTT, but not stress-induced DoG transcription, was accompanied by a strong increase in open chromatin downstream of the affected poly(A) sites. In its extent and kinetics, downstream open chromatin essentially matched the poly(A) read-through transcription. We show that this does not cause but rather requires DoTT as well as high levels of transcription into the genomic regions downstream of genes. This raises intriguing new questions regarding the role of histone repositioning in the wake of RNA Polymerase II passage downstream of impaired poly(A) site recognition.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Herpes Simplex Virus 1 Dramatically Alters Loading and Positioning of RNA Polymerase II on Host Genes Early in Infection.J Virol. 2018 Mar 28;92(8):e02184-17. doi: 10.1128/JVI.02184-17. Print 2018 Apr 15. J Virol. 2018. PMID: 29437966 Free PMC article.
-
Widespread disruption of host transcription termination in HSV-1 infection.Nat Commun. 2015 May 20;6:7126. doi: 10.1038/ncomms8126. Nat Commun. 2015. PMID: 25989971 Free PMC article.
-
The HSV-1 ICP22 protein selectively impairs histone repositioning upon Pol II transcription downstream of genes.Nat Commun. 2023 Jul 31;14(1):4591. doi: 10.1038/s41467-023-40217-w. Nat Commun. 2023. PMID: 37524699 Free PMC article.
-
Computational Integration of HSV-1 Multi-omics Data.Methods Mol Biol. 2023;2610:31-48. doi: 10.1007/978-1-0716-2895-9_3. Methods Mol Biol. 2023. PMID: 36534279 Review.
-
Chromatin dynamics during herpes simplex virus-1 lytic infection.Biochim Biophys Acta. 2010 Mar-Apr;1799(3-4):223-7. doi: 10.1016/j.bbagrm.2010.01.012. Epub 2010 Feb 6. Biochim Biophys Acta. 2010. PMID: 20139038 Review.
Cited by
-
Dynamic chromatin accessibility profiling reveals changes in host genome organization in response to baculovirus infection.PLoS Pathog. 2020 Jun 8;16(6):e1008633. doi: 10.1371/journal.ppat.1008633. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32511266 Free PMC article.
-
Mechanisms and consequences of mRNA destabilization during viral infections.Virol J. 2024 Feb 6;21(1):38. doi: 10.1186/s12985-024-02305-1. Virol J. 2024. PMID: 38321453 Free PMC article. Review.
-
Herpes simplex virus 1 inhibits phosphorylation of RNA polymerase II CTD serine-7.J Virol. 2024 Oct 22;98(10):e0117824. doi: 10.1128/jvi.01178-24. Epub 2024 Sep 24. J Virol. 2024. PMID: 39316591 Free PMC article.
-
Characterization of influenza A virus induced transposons reveals a subgroup of transposons likely possessing the regulatory role as eRNAs.Sci Rep. 2022 Feb 9;12(1):2188. doi: 10.1038/s41598-022-06196-6. Sci Rep. 2022. PMID: 35140280 Free PMC article.
-
Mechanisms of lncRNA biogenesis as revealed by nascent transcriptomics.Nat Rev Mol Cell Biol. 2022 Jun;23(6):389-406. doi: 10.1038/s41580-021-00447-6. Epub 2022 Jan 25. Nat Rev Mol Cell Biol. 2022. PMID: 35079163 Review.
References
-
- Porrua O, Libri D. Transcription termination and the control of the transcriptome: why, where and how to stop. Nat Rev Mol Cell Biol. 2015;16(3):190–202. doi: 10.1038/nrm3943 - DOI - PubMed
-
- Proudfoot NJ. Transcriptional termination in mammals: Stopping the RNA polymerase II juggernaut. Science. 2016;352(6291):aad9926 doi: 10.1126/science.aad9926 ; PubMed Central PMCID: PMCPMC5144996. - DOI - PMC - PubMed
-
- Sandri-Goldin RM. The many roles of the regulatory protein ICP27 during herpes simplex virus infection. Front Biosci. 2008;13:5241–56. Epub 2008/05/30. doi: https://doi.org/10.2741/3078 [pii]. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
