

CD150high Bone Marrow Tregs Maintain Hematopoietic Stem Cell Quiescence and Immune Privilege via Adenosine
- PMID: 29456159
- PMCID: PMC6534147
- DOI: 10.1016/j.stem.2018.01.017
CD150high Bone Marrow Tregs Maintain Hematopoietic Stem Cell Quiescence and Immune Privilege via Adenosine
Abstract
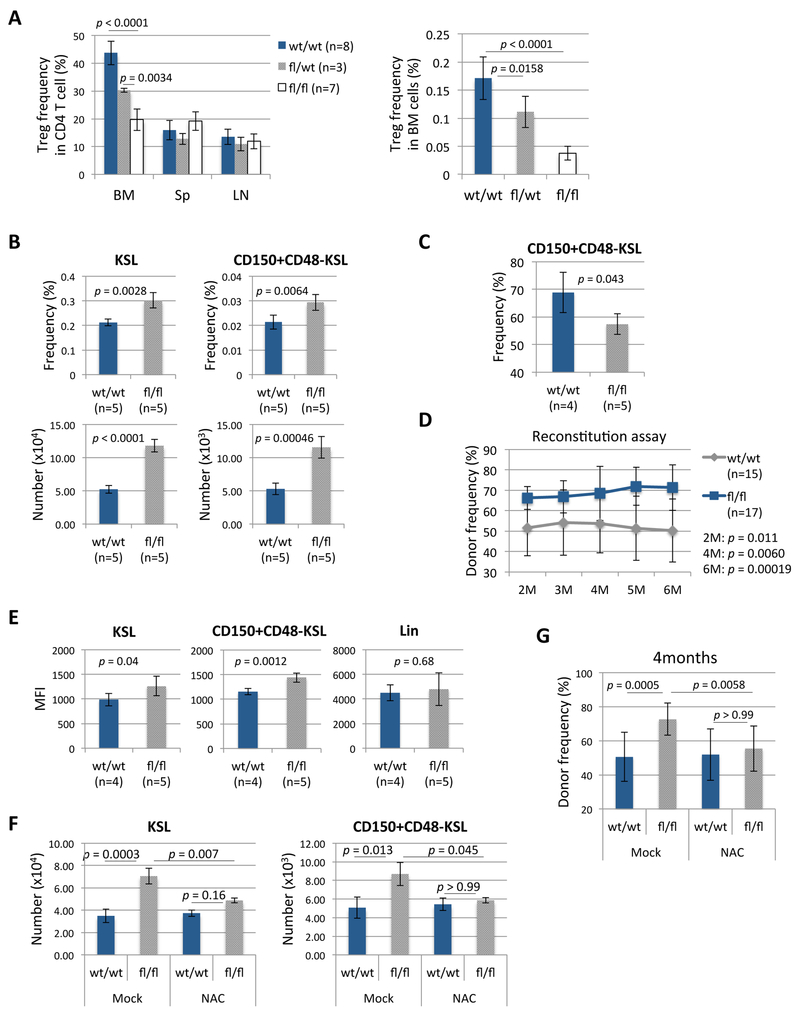
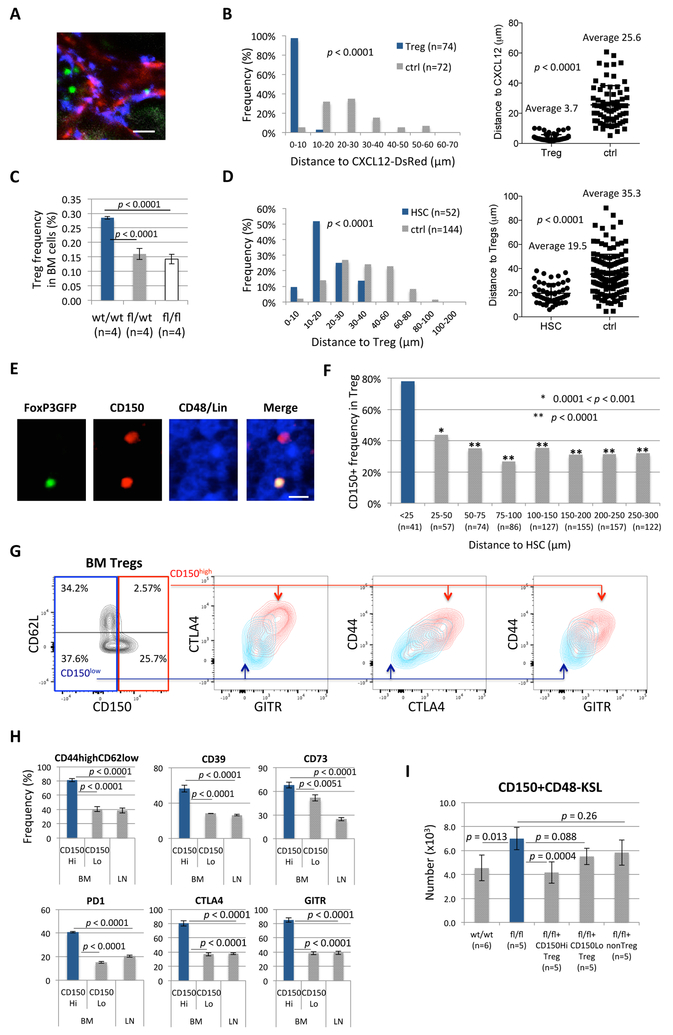
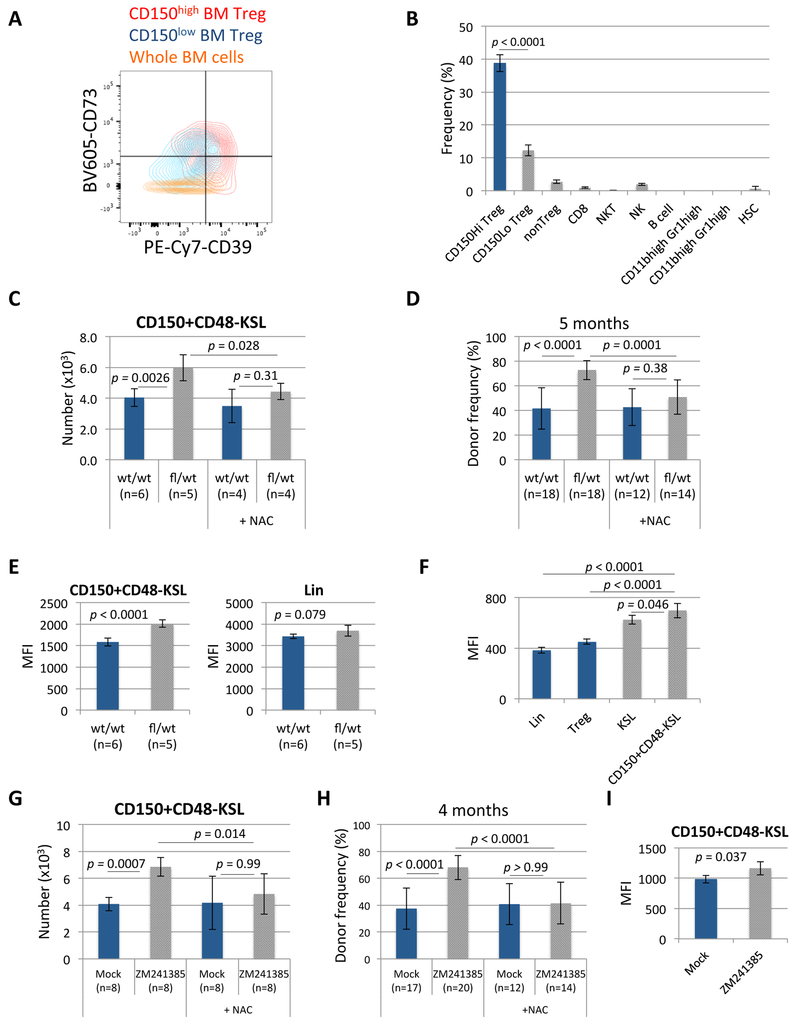
A crucial player in immune regulation, FoxP3+ regulatory T cells (Tregs) are drawing attention for their heterogeneity and noncanonical functions. Here, we describe a Treg subpopulation that controls hematopoietic stem cell (HSC) quiescence and engraftment. These Tregs highly expressed an HSC marker, CD150, and localized within the HSC niche in the bone marrow (BM). Specific reduction of BM Tregs achieved by conditional deletion of CXCR4 in Tregs increased HSC numbers in the BM. Adenosine generated via the CD39 cell surface ectoenzyme on niche Tregs protected HSCs from oxidative stress and maintained HSC quiescence. In transplantation settings, niche Tregs prevented allogeneic (allo-) HSC rejection through adenosine and facilitated allo-HSC engraftment. Furthermore, transfer of niche Tregs promoted allo-HSC engraftment to a much greater extent than transfer of other Tregs. These results identify a unique niche-associated Treg subset and adenosine as regulators of HSC quiescence, abundance, and engraftment, further highlighting their therapeutic utility.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors have declared that no conflicting interests exist.
Figures

Similar articles
-
CD150high CD4 T cells and CD150high regulatory T cells regulate hematopoietic stem cell quiescence via CD73.Haematologica. 2019 Jun;104(6):1136-1142. doi: 10.3324/haematol.2018.198283. Epub 2018 Dec 13. Haematologica. 2019. PMID: 30545927 Free PMC article.
-
Paradoxical Regulation of Allogeneic Bone Marrow Engraftment and Immune Privilege by Mesenchymal Cells and Adenosine.Transplant Cell Ther. 2021 Jan;27(1):92.e1-92.e5. doi: 10.1016/j.bbmt.2020.09.017. Epub 2020 Sep 19. Transplant Cell Ther. 2021. PMID: 32961376 Free PMC article.
-
CD166 regulates human and murine hematopoietic stem cells and the hematopoietic niche.Blood. 2014 Jul 24;124(4):519-29. doi: 10.1182/blood-2014-03-565721. Epub 2014 Apr 16. Blood. 2014. PMID: 24740813 Free PMC article.
-
Regulation of hematopoietic and leukemia stem cells by regulatory T cells.Front Immunol. 2022 Nov 2;13:1049301. doi: 10.3389/fimmu.2022.1049301. eCollection 2022. Front Immunol. 2022. PMID: 36405718 Free PMC article. Review.
-
Regulatory T cell niche in the bone marrow, a new player in Haematopoietic stem cell transplantation.Blood Rev. 2023 May;59:101030. doi: 10.1016/j.blre.2022.101030. Epub 2022 Oct 31. Blood Rev. 2023. PMID: 36336520 Review.
Cited by
-
PD-L1 blockade immunotherapy rewires cancer-induced emergency myelopoiesis.Front Immunol. 2024 Oct 11;15:1386838. doi: 10.3389/fimmu.2024.1386838. eCollection 2024. Front Immunol. 2024. PMID: 39464894 Free PMC article.
-
Bone Marrow Niches of Hematopoietic Stem and Progenitor Cells.Int J Mol Sci. 2022 Apr 18;23(8):4462. doi: 10.3390/ijms23084462. Int J Mol Sci. 2022. PMID: 35457280 Free PMC article. Review.
-
Three-Dimensional Avian Hematopoietic Stem Cell Cultures as a Model for Studying Disease Pathogenesis.Front Cell Dev Biol. 2022 Jan 20;9:730804. doi: 10.3389/fcell.2021.730804. eCollection 2021. Front Cell Dev Biol. 2022. PMID: 35127695 Free PMC article. Review.
-
CD150high CD4 T cells and CD150high regulatory T cells regulate hematopoietic stem cell quiescence via CD73.Haematologica. 2019 Jun;104(6):1136-1142. doi: 10.3324/haematol.2018.198283. Epub 2018 Dec 13. Haematologica. 2019. PMID: 30545927 Free PMC article.
-
Tnfrsf4-expressing regulatory T cells promote immune escape of chronic myeloid leukemia stem cells.JCI Insight. 2021 Dec 8;6(23):e151797. doi: 10.1172/jci.insight.151797. JCI Insight. 2021. PMID: 34727093 Free PMC article.
References
-
- Azab AK, Runnels JM, Pitsillides C, Moreau AS, Azab F, Leleu X, Jia X, Wright R, Ospina B, Carlson AL, et al. (2009). CXCR4 inhibitor AMD3100 disrupts the interaction of multiple myeloma cells with the bone marrow microenvironment and enhances their sensitivity to therapy. Blood, 113, 4341–51. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
