

iRhom2 promotes lupus nephritis through TNF-α and EGFR signaling
- PMID: 29369823
- PMCID: PMC5873859
- DOI: 10.1172/JCI97650
iRhom2 promotes lupus nephritis through TNF-α and EGFR signaling
Abstract
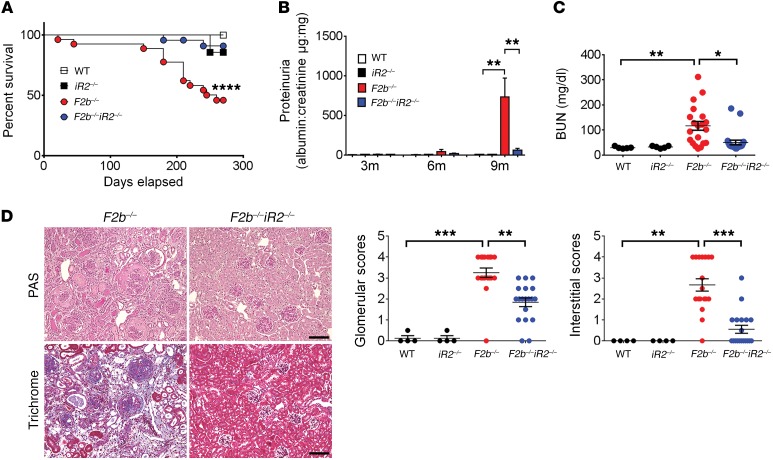
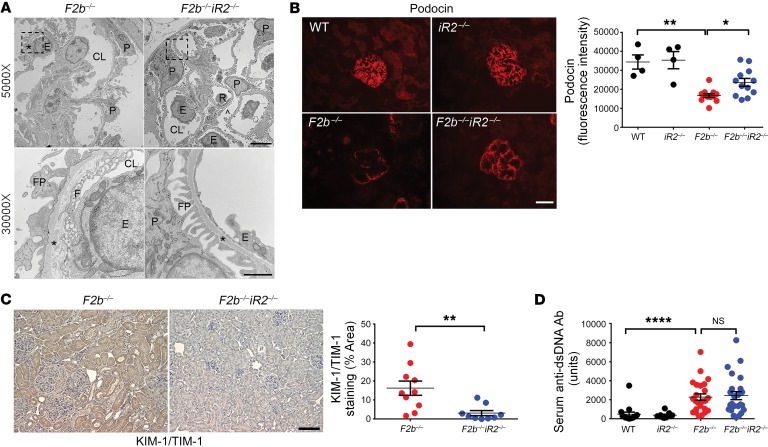
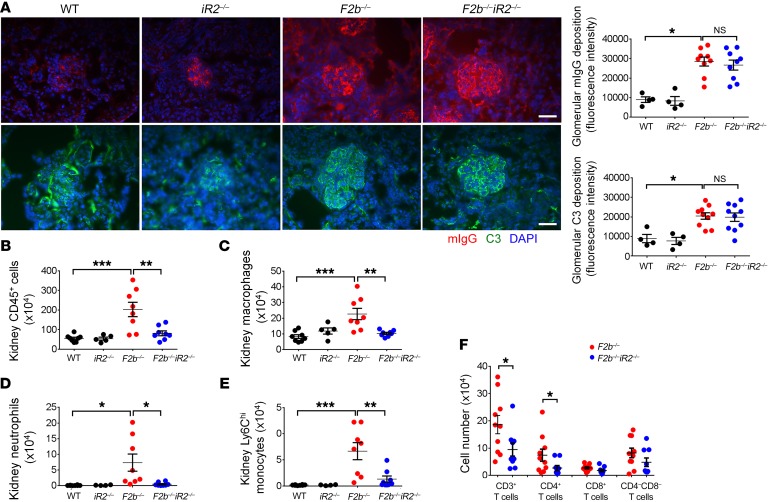
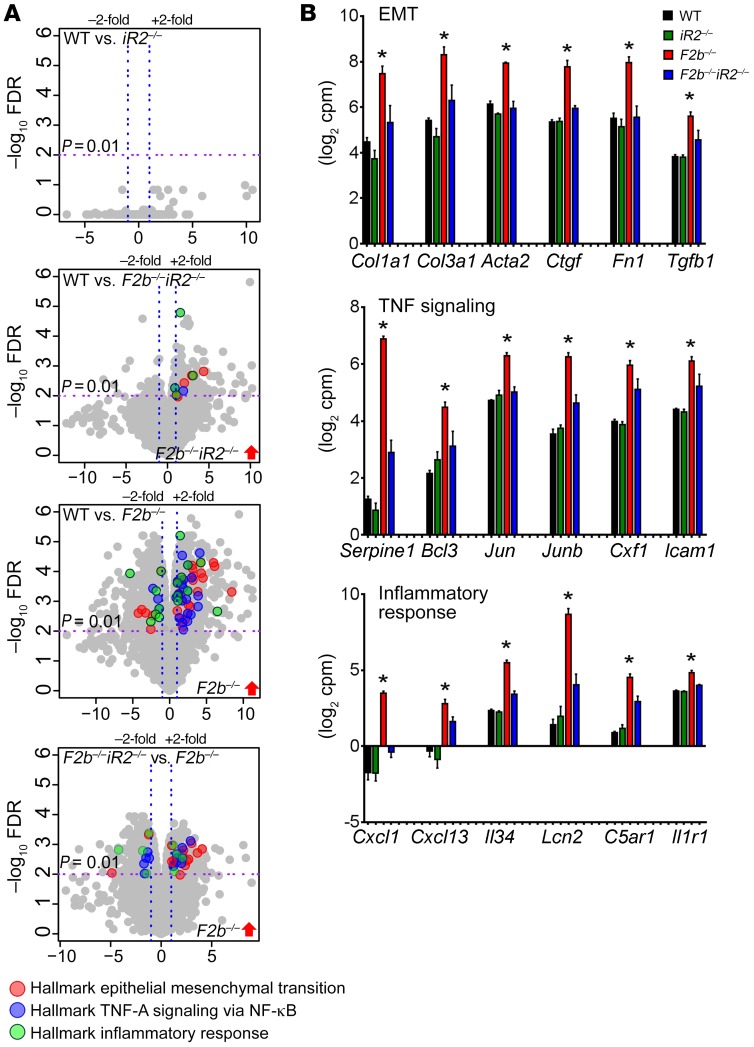
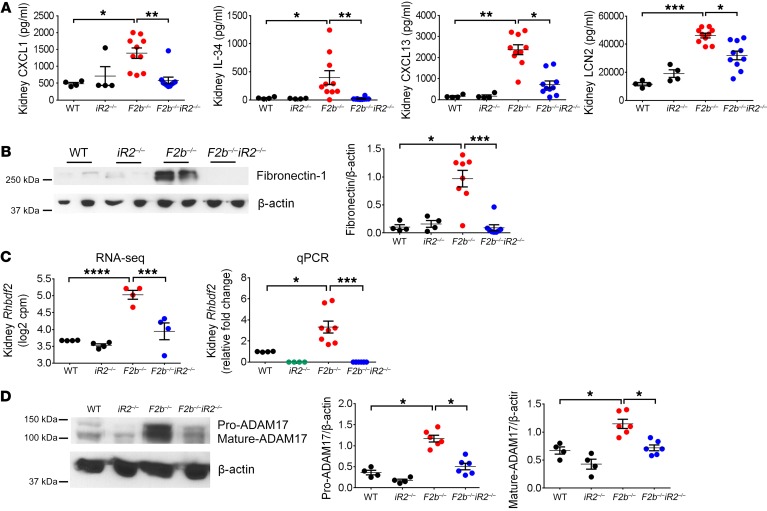
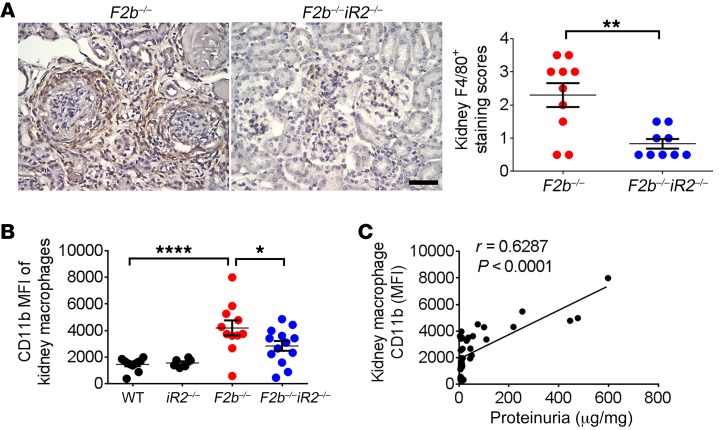
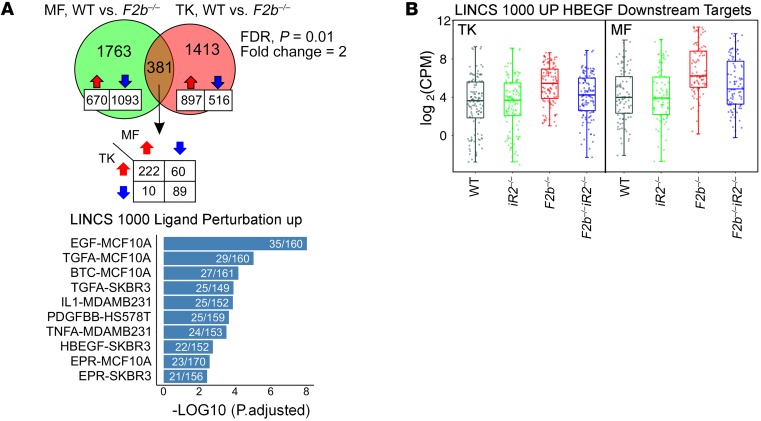
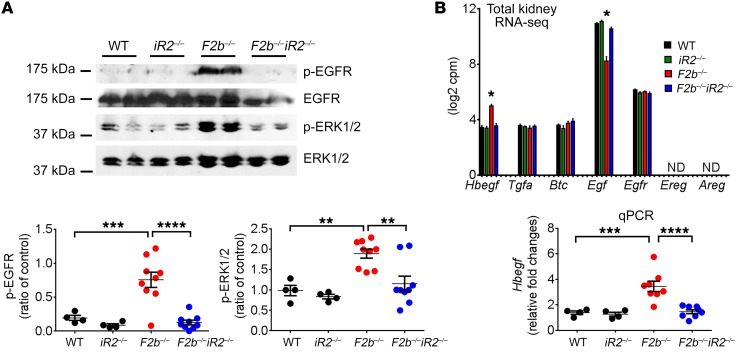
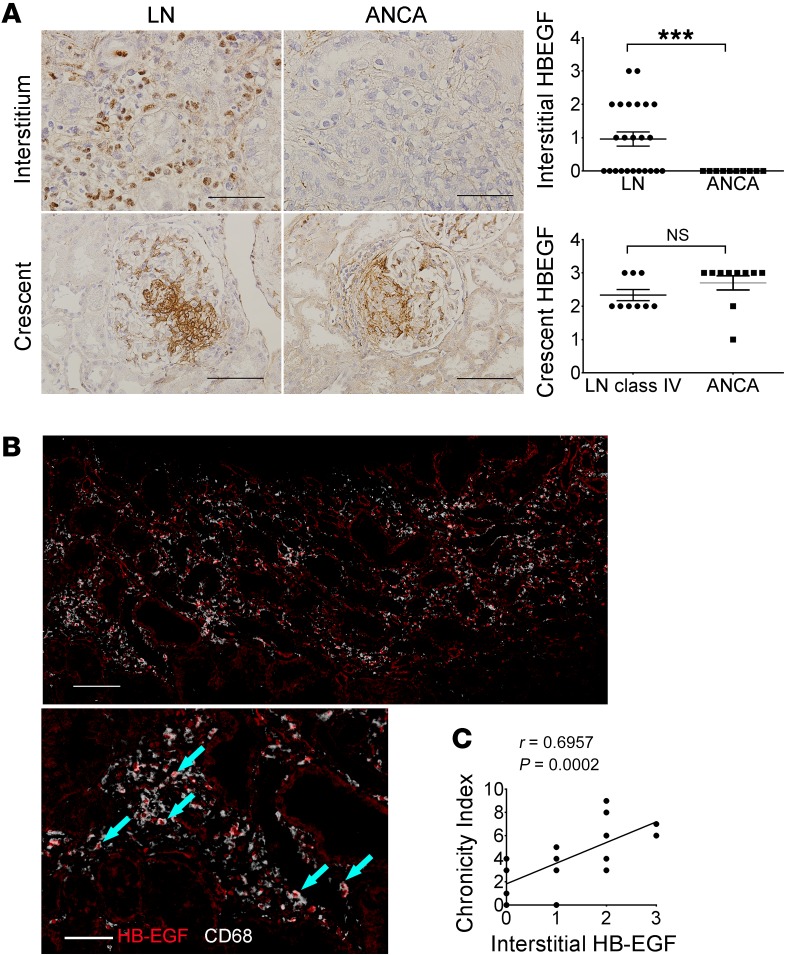
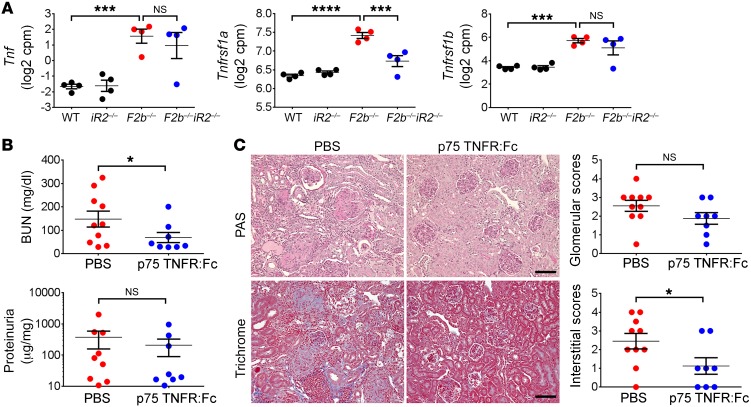
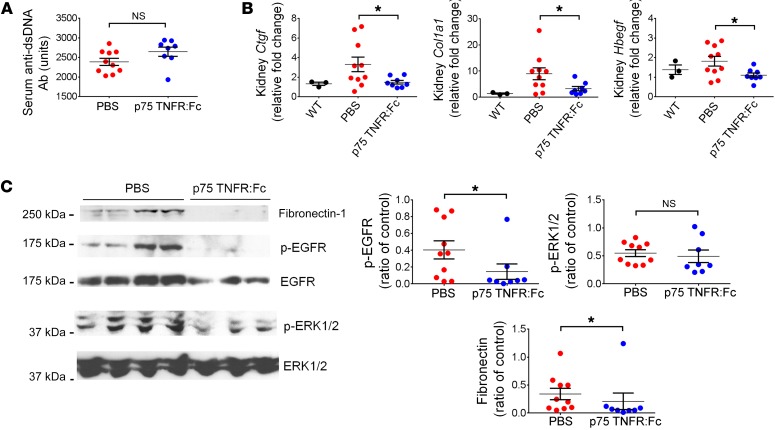
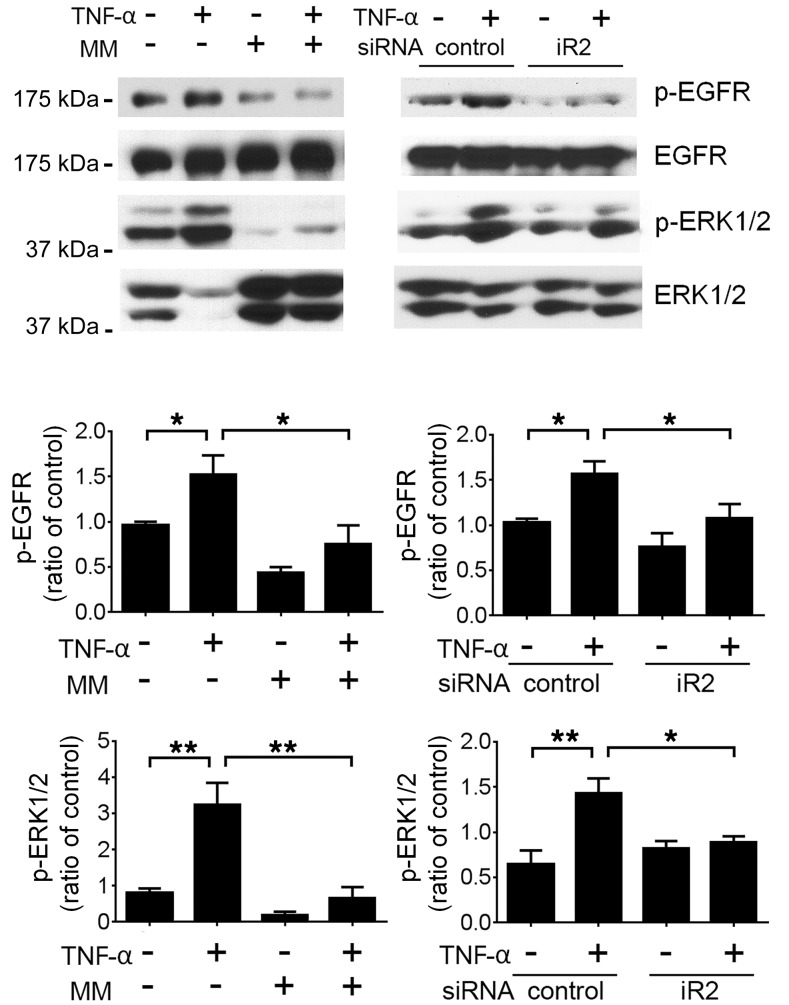
Lupus nephritis (LN) often results in progressive renal dysfunction. The inactive rhomboid 2 (iRhom2) is a newly identified key regulator of A disintegrin and metalloprotease 17 (ADAM17), whose substrates, such as TNF-α and heparin-binding EGF (HB-EGF), have been implicated in the pathogenesis of chronic kidney diseases. Here, we demonstrate that deficiency of iRhom2 protects the lupus-prone Fcgr2b-/- mice from developing severe kidney damage without altering anti-double-stranded DNA (anti-dsDNA) Ab production by simultaneously blocking HB-EGF/EGFR and TNF-α signaling in the kidney tissues. Unbiased transcriptome profiling of kidneys and kidney macrophages revealed that TNF-α and HB-EGF/EGFR signaling pathways are highly upregulated in Fcgr2b-/- mice, alterations that were diminished in the absence of iRhom2. Pharmacological blockade of either TNF-α or EGFR signaling protected Fcgr2b-/- mice from severe renal damage. Finally, kidneys from LN patients showed increased iRhom2 and HB-EGF expression, with interstitial HB-EGF expression significantly associated with chronicity indices. Our data suggest that activation of iRhom2/ADAM17-dependent TNF-α and EGFR signaling plays a crucial role in mediating irreversible kidney damage in LN, thereby uncovering a target for selective and simultaneous dual inhibition of 2 major pathological pathways in the effector arm of the disease.
Keywords: Autoimmunity; Inflammation; Lupus; Mouse models.
Conflict of interest statement
Figures

Comment in
-
iRhoms: A Potential Path to More Specific Therapeutic Targeting of Lupus Nephritis.Am J Kidney Dis. 2018 Oct;72(4):617-619. doi: 10.1053/j.ajkd.2018.04.010. Epub 2018 Jun 7. Am J Kidney Dis. 2018. PMID: 29887489 No abstract available.
Similar articles
-
The xenoestrogens biphenol-A and nonylphenol differentially regulate metalloprotease-mediated shedding of EGFR ligands.J Cell Physiol. 2018 Mar;233(3):2247-2256. doi: 10.1002/jcp.26097. Epub 2017 Aug 25. J Cell Physiol. 2018. PMID: 28703301 Free PMC article.
-
Activated iRhom2 drives prolonged PM2.5 exposure-triggered renal injury in Nrf2-defective mice.Nanotoxicology. 2018 Nov;12(9):1045-1067. doi: 10.1080/17435390.2018.1513093. Epub 2018 Sep 26. Nanotoxicology. 2018. PMID: 30257117
-
ADAM17 stabilizes its interacting partner inactive Rhomboid 2 (iRhom2) but not inactive Rhomboid 1 (iRhom1).J Biol Chem. 2020 Mar 27;295(13):4350-4358. doi: 10.1074/jbc.RA119.011136. Epub 2020 Feb 14. J Biol Chem. 2020. PMID: 32060096 Free PMC article.
-
iRhom2: An Emerging Adaptor Regulating Immunity and Disease.Int J Mol Sci. 2020 Sep 8;21(18):6570. doi: 10.3390/ijms21186570. Int J Mol Sci. 2020. PMID: 32911849 Free PMC article. Review.
-
The Role of iRhom2 in Metabolic and Cardiovascular-Related Disorders.Front Cardiovasc Med. 2020 Nov 24;7:612808. doi: 10.3389/fcvm.2020.612808. eCollection 2020. Front Cardiovasc Med. 2020. PMID: 33330676 Free PMC article. Review.
Cited by
-
Targeting the epidermal growth factor receptor (EGFR/ErbB) for the potential treatment of renal pathologies.Front Pharmacol. 2024 Aug 21;15:1394997. doi: 10.3389/fphar.2024.1394997. eCollection 2024. Front Pharmacol. 2024. PMID: 39234105 Free PMC article. Review.
-
iRhom2 deficiency reduces sepsis-induced mortality associated with the attenuation of lung macrophages in mice.Histochem Cell Biol. 2024 Nov;162(5):415-428. doi: 10.1007/s00418-024-02318-5. Epub 2024 Aug 12. Histochem Cell Biol. 2024. PMID: 39134731 Free PMC article.
-
The interferon-rich skin environment regulates Langerhans cell ADAM17 to promote photosensitivity in lupus.Elife. 2024 Jun 11;13:e85914. doi: 10.7554/eLife.85914. Elife. 2024. PMID: 38860651 Free PMC article.
-
Th1-related transcription factors and cytokines in systemic lupus erythematosus.Front Immunol. 2023 Dec 18;14:1305590. doi: 10.3389/fimmu.2023.1305590. eCollection 2023. Front Immunol. 2023. PMID: 38164134 Free PMC article. Review.
-
Epidermal growth factor receptor activation is essential for kidney fibrosis development.Nat Commun. 2023 Nov 14;14(1):7357. doi: 10.1038/s41467-023-43226-x. Nat Commun. 2023. PMID: 37963889 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
