

Dual Strategies for Argonaute2-Mediated Biogenesis of Erythroid miRNAs Underlie Conserved Requirements for Slicing in Mammals
- PMID: 29351846
- PMCID: PMC5824974
- DOI: 10.1016/j.molcel.2017.12.027
Dual Strategies for Argonaute2-Mediated Biogenesis of Erythroid miRNAs Underlie Conserved Requirements for Slicing in Mammals
Abstract
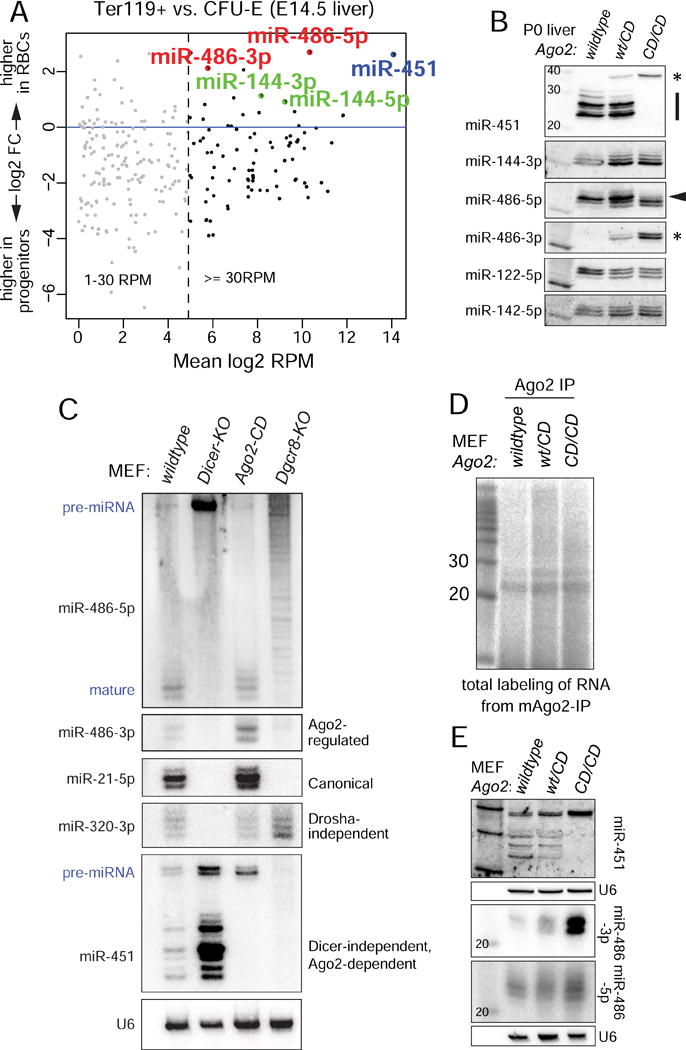
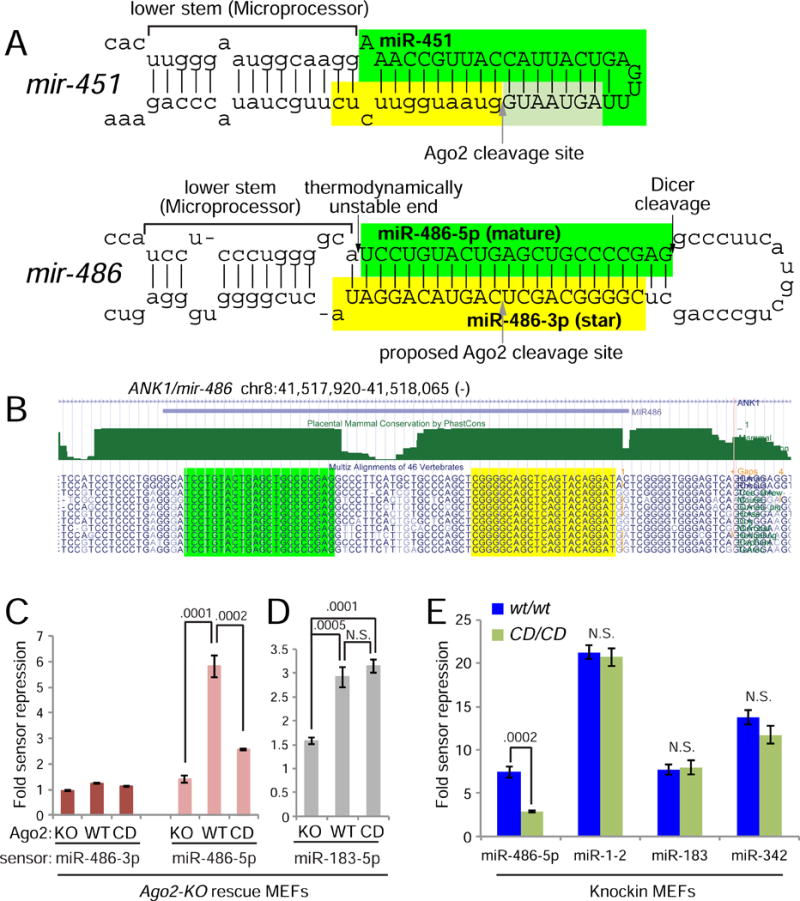
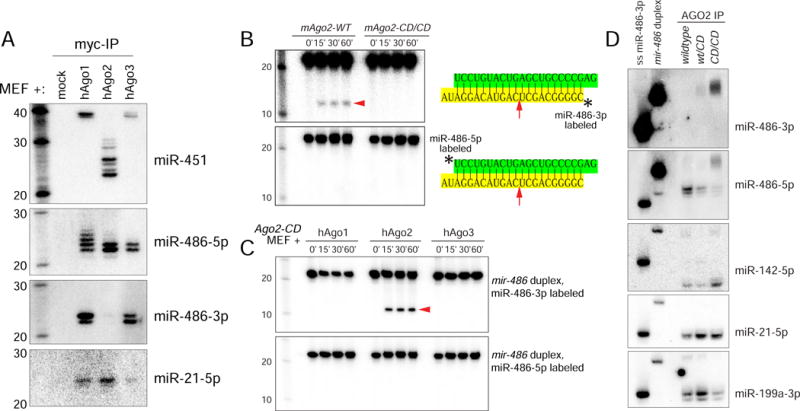
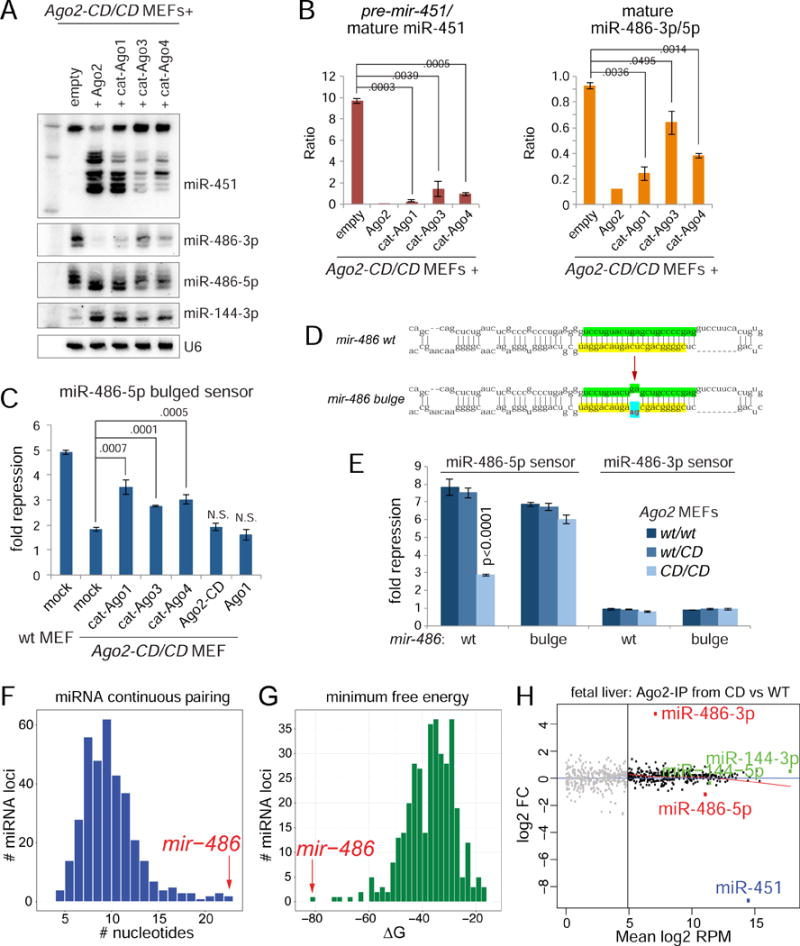
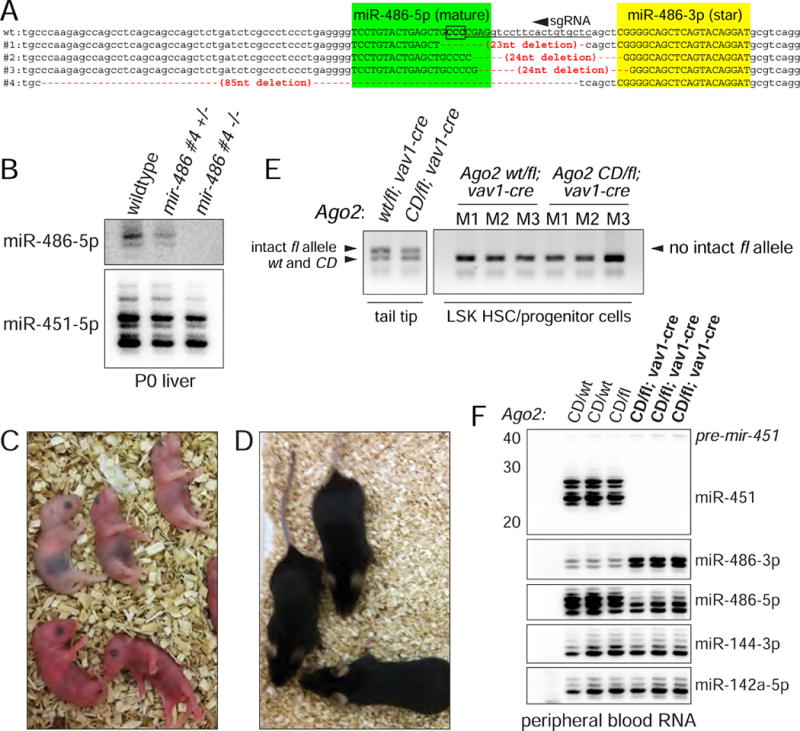
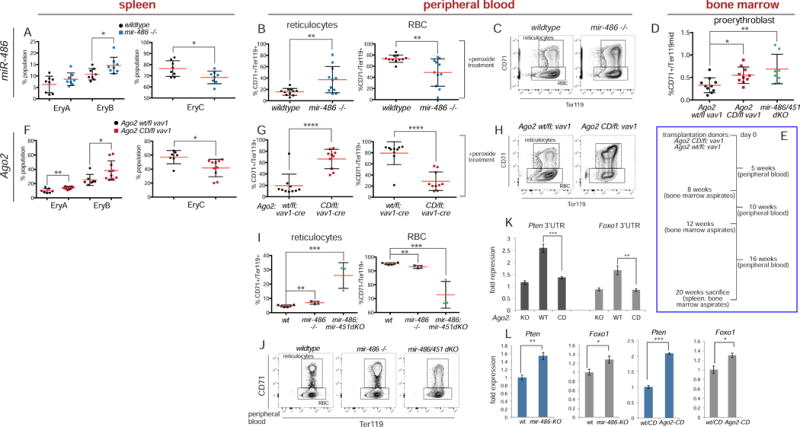
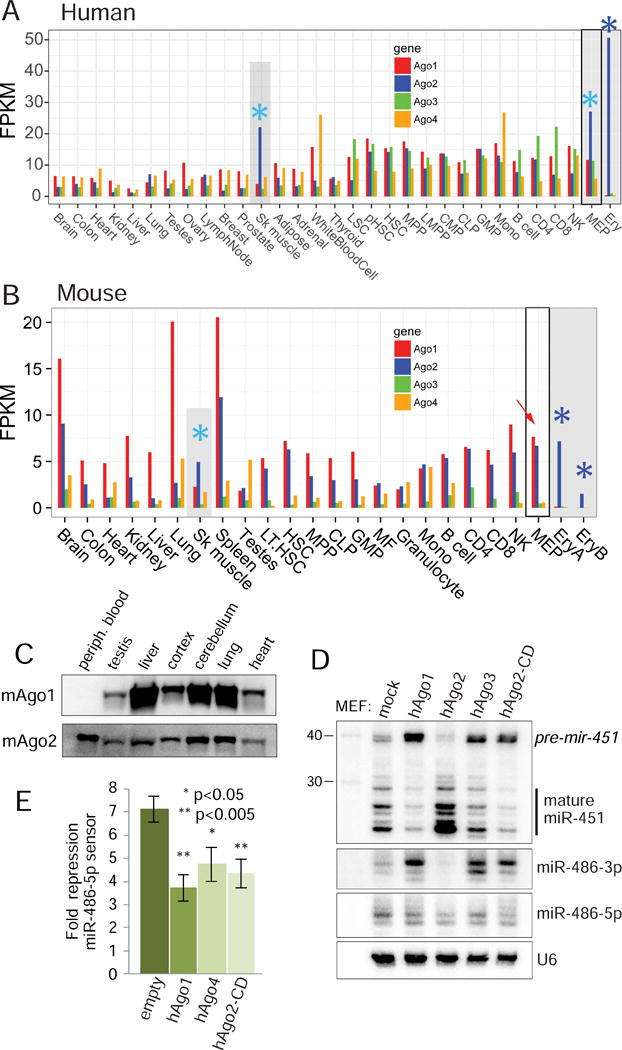
While Slicer activity of Argonaute is central to RNAi, conserved roles of slicing in endogenous regulatory biology are less clear, especially in mammals. Biogenesis of erythroid Dicer-independent mir-451 involves Ago2 catalysis, but mir-451-KO mice do not phenocopy Ago2 catalytic-dead (Ago2-CD) mice, suggesting other needs for slicing. Here, we reveal mir-486 as another dominant erythroid miRNA with atypical biogenesis. While it is Dicer dependent, it requires slicing to eliminate its star strand. Thus, in Ago2-CD conditions, miR-486-5p is functionally inactive due to duplex arrest. Genome-wide analyses reveal miR-486 and miR-451 as the major slicing-dependent miRNAs in the hematopoietic system. Moreover, mir-486-KO mice exhibit erythroid defects, and double knockout of mir-486/451 phenocopies the cell-autonomous effects of Ago2-CD in the hematopoietic system. Finally, we observe that Ago2 is the dominant-expressed Argonaute in maturing erythroblasts, reflecting a specialized environment for processing slicing-dependent miRNAs. Overall, the mammalian hematopoietic system has evolved multiple conserved requirements for Slicer-dependent miRNA biogenesis.
Keywords: Ago2; Argonaute; RNAi; Slicer; erythropoiesis; hematopoietic; miR-451; miR-486; miRNA; microRNA.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
There are no competing interests.
Figures
Similar articles
-
Ago2-Dependent Processing Allows miR-451 to Evade the Global MicroRNA Turnover Elicited during Erythropoiesis.Mol Cell. 2020 Apr 16;78(2):317-328.e6. doi: 10.1016/j.molcel.2020.02.020. Epub 2020 Mar 18. Mol Cell. 2020. PMID: 32191872 Free PMC article.
-
Conserved vertebrate mir-451 provides a platform for Dicer-independent, Ago2-mediated microRNA biogenesis.Proc Natl Acad Sci U S A. 2010 Aug 24;107(34):15163-8. doi: 10.1073/pnas.1006432107. Epub 2010 Aug 10. Proc Natl Acad Sci U S A. 2010. PMID: 20699384 Free PMC article.
-
eIF1A augments Ago2-mediated Dicer-independent miRNA biogenesis and RNA interference.Nat Commun. 2015 May 28;6:7194. doi: 10.1038/ncomms8194. Nat Commun. 2015. PMID: 26018492 Free PMC article.
-
Dicer-independent processing of small RNA duplexes: mechanistic insights and applications.Nucleic Acids Res. 2017 Oct 13;45(18):10369-10379. doi: 10.1093/nar/gkx779. Nucleic Acids Res. 2017. PMID: 28977573 Free PMC article. Review.
-
New insights into the function of mammalian Argonaute2.PLoS Genet. 2020 Nov 12;16(11):e1009058. doi: 10.1371/journal.pgen.1009058. eCollection 2020 Nov. PLoS Genet. 2020. PMID: 33180792 Free PMC article. Review.
Cited by
-
Global Switch from DICER-dependent MicroRNA to DICER-independent SnoRNA-derived RNA Biogenesis in Malignancy.MicroPubl Biol. 2023 Feb 2;2023:10.17912/micropub.biology.000725. doi: 10.17912/micropub.biology.000725. eCollection 2023. MicroPubl Biol. 2023. PMID: 36818311 Free PMC article.
-
Parameters of clustered suboptimal miRNA biogenesis.Proc Natl Acad Sci U S A. 2023 Oct 10;120(41):e2306727120. doi: 10.1073/pnas.2306727120. Epub 2023 Oct 3. Proc Natl Acad Sci U S A. 2023. PMID: 37788316 Free PMC article.
-
Depletion of erythropoietic miR-486-5p and miR-451a improves detectability of rare microRNAs in peripheral blood-derived small RNA sequencing libraries.NAR Genom Bioinform. 2020 Feb 12;2(1):lqaa008. doi: 10.1093/nargab/lqaa008. eCollection 2020 Mar. NAR Genom Bioinform. 2020. PMID: 33575555 Free PMC article.
-
The importance of cellular and exosomal miRNAs in mesenchymal stem cell osteoblastic differentiation.Sci Rep. 2021 Mar 15;11(1):5953. doi: 10.1038/s41598-021-85306-2. Sci Rep. 2021. PMID: 33723364 Free PMC article.
-
Identification of miR-18a-5p as an oncogene and prognostic biomarker in RCC.Am J Transl Res. 2018 Jun 15;10(6):1874-1886. eCollection 2018. Am J Transl Res. 2018. PMID: 30018727 Free PMC article.
References
-
- Alexander MS, Casar JC, Motohashi N, Vieira NM, Eisenberg I, Marshall JL, Gasperini MJ, Lek A, Myers JA, Estrella EA, et al. MicroRNA-486-dependent modulation of DOCK3/PTEN/AKT signaling pathways improves muscular dystrophy-associated symptoms. The Journal of clinical investigation. 2014;124:2651–2667. - PMC - PubMed
-
- Bianchi E, Bulgarelli J, Ruberti S, Rontauroli S, Sacchi G, Norfo R, Pennucci V, Zini R, Salati S, Prudente Z, et al. MYB controls erythroid versus megakaryocyte lineage fate decision through the miR-486-3p-mediated downregulation of MAF. Cell death and differentiation. 2015;22:1906–1921. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
