

Spatiotemporal allele organization by allele-specific CRISPR live-cell imaging (SNP-CLING)
- PMID: 29343869
- PMCID: PMC5805655
- DOI: 10.1038/s41594-017-0015-3
Spatiotemporal allele organization by allele-specific CRISPR live-cell imaging (SNP-CLING)
Abstract
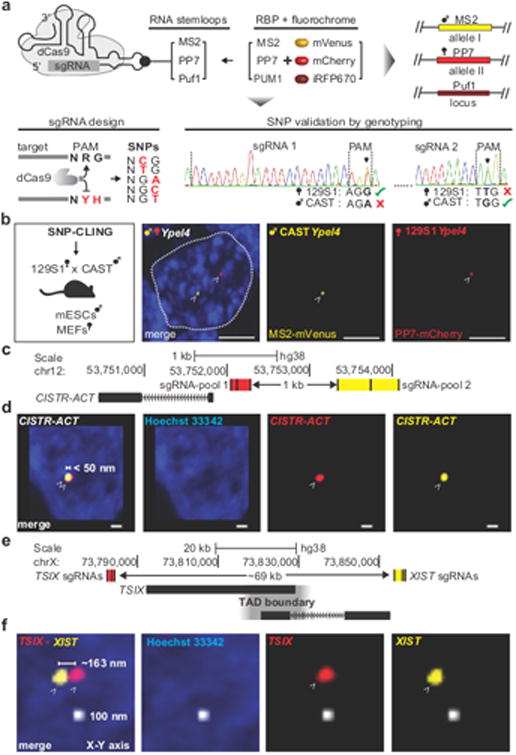
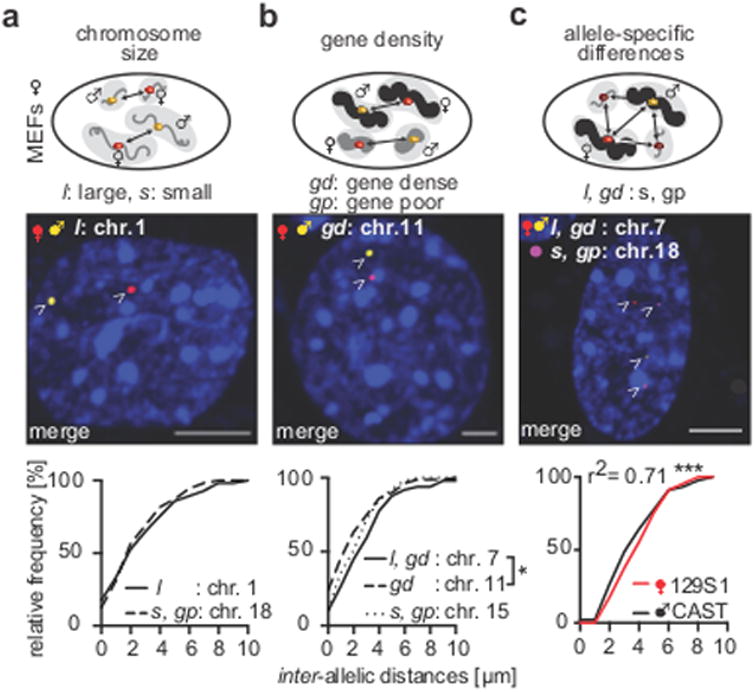
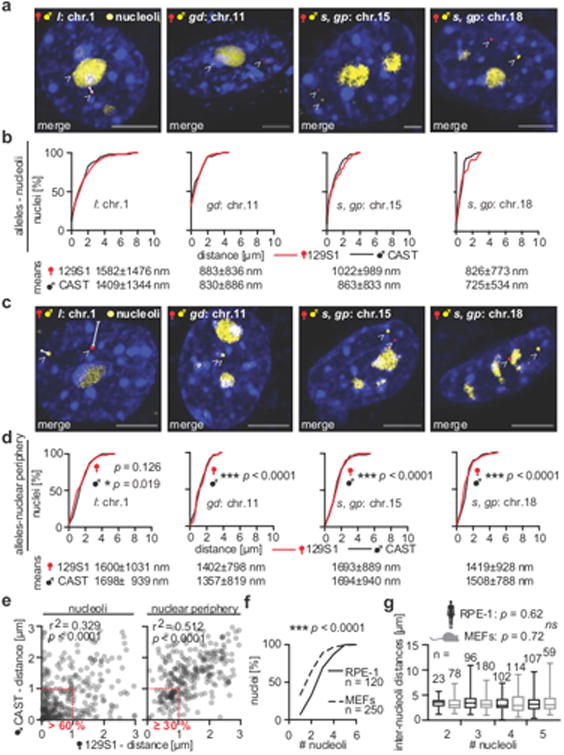
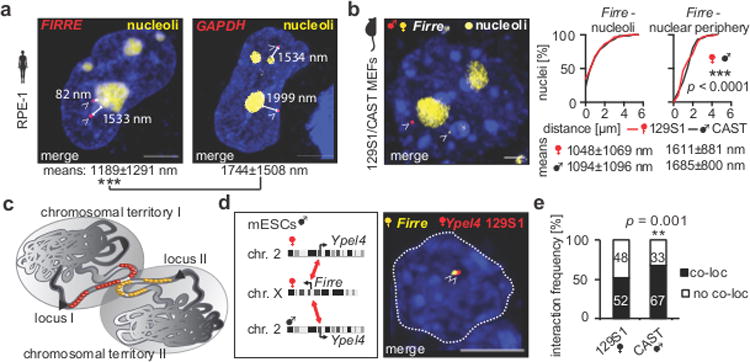
Imaging and chromatin capture techniques have provided important insights into our understanding of nuclear organization. A limitation of these techniques is the inability to resolve allele-specific spatiotemporal properties of genomic loci in living cells. Here, we describe an allele-specific CRISPR live-cell DNA imaging technique (SNP-CLING) to provide the first comprehensive insights into allelic positioning across space and time in mouse embryonic stem cells and fibroblasts. With 3D imaging, we studied alleles on different chromosomes in relation to one another and relative to nuclear substructures such as the nucleolus. We find that alleles maintain similar positions relative to each other and the nucleolus; however, loci occupy unique positions. To monitor spatiotemporal dynamics by SNP-CLING, we performed 4D imaging and determined that alleles are either stably positioned or fluctuating during cell state transitions, such as apoptosis. SNP-CLING is a universally applicable technique that enables the dissection of allele-specific spatiotemporal genome organization in live cells.
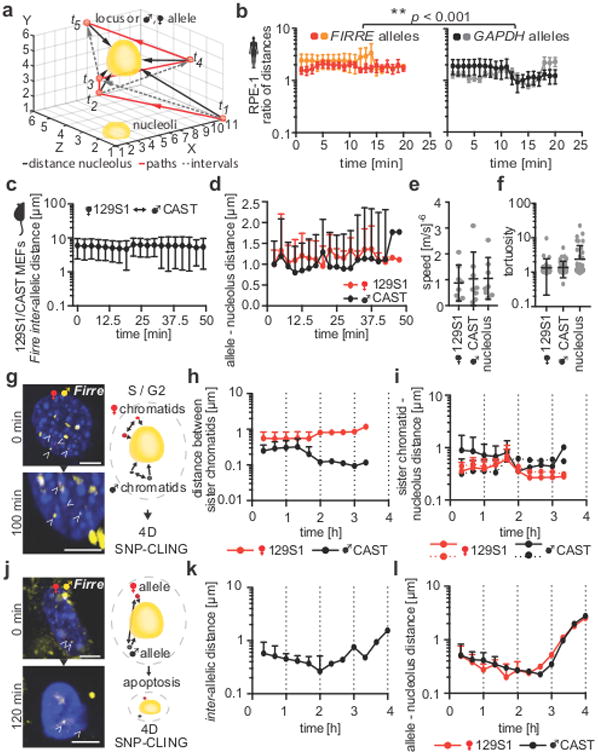
Figures
Comment in
-
Technique: SNP-CLINGing onto your post in the genome.Nat Rev Genet. 2018 Mar;19(3):126-127. doi: 10.1038/nrg.2018.2. Epub 2018 Jan 22. Nat Rev Genet. 2018. PMID: 29353874 No abstract available.
Similar articles
-
Multiplexed Dynamic Imaging of Genomic Loci by Combined CRISPR Imaging and DNA Sequential FISH.Biophys J. 2017 May 9;112(9):1773-1776. doi: 10.1016/j.bpj.2017.03.024. Epub 2017 Apr 17. Biophys J. 2017. PMID: 28427715 Free PMC article.
-
CRISPR-based genomic loci labeling revealed ordered spatial organization of chromatin in living diploid human cells.Biochim Biophys Acta Mol Cell Res. 2019 Dec;1866(12):118518. doi: 10.1016/j.bbamcr.2019.07.013. Epub 2019 Jul 31. Biochim Biophys Acta Mol Cell Res. 2019. PMID: 31376419
-
Inter-chromosomal Contact Properties in Live-Cell Imaging and in Hi-C.Mol Cell. 2018 Mar 15;69(6):1039-1045.e3. doi: 10.1016/j.molcel.2018.02.007. Epub 2018 Mar 8. Mol Cell. 2018. PMID: 29526697 Free PMC article.
-
[Advances in application of clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated 9 system in stem cells research].Zhonghua Shao Shang Za Zhi. 2018 Apr 20;34(4):253-256. doi: 10.3760/cma.j.issn.1009-2587.2018.04.013. Zhonghua Shao Shang Za Zhi. 2018. PMID: 29690746 Review. Chinese.
-
Imaging Specific Genomic DNA in Living Cells.Annu Rev Biophys. 2016 Jul 5;45:1-23. doi: 10.1146/annurev-biophys-062215-010830. Epub 2016 Apr 27. Annu Rev Biophys. 2016. PMID: 27145877 Free PMC article. Review.
Cited by
-
Next-generation CRISPR technology for genome, epigenome and mitochondrial editing.Transgenic Res. 2024 Oct;33(5):323-357. doi: 10.1007/s11248-024-00404-x. Epub 2024 Aug 19. Transgenic Res. 2024. PMID: 39158822 Review.
-
Recent advances of the biological and biomedical applications of CRISPR/Cas systems.Mol Biol Rep. 2022 Jul;49(7):7087-7100. doi: 10.1007/s11033-022-07519-6. Epub 2022 Jun 15. Mol Biol Rep. 2022. PMID: 35705772 Free PMC article. Review.
-
In vivo Firre and Dxz4 deletion elucidates roles for autosomal gene regulation.Elife. 2019 Nov 18;8:e47214. doi: 10.7554/eLife.47214. Elife. 2019. PMID: 31738164 Free PMC article.
-
Diversification of the CRISPR Toolbox: Applications of CRISPR-Cas Systems Beyond Genome Editing.CRISPR J. 2021 Jun;4(3):400-415. doi: 10.1089/crispr.2020.0137. CRISPR J. 2021. PMID: 34152221 Free PMC article.
-
CRISPR-Cas12a Nucleases Bind Flexible DNA Duplexes without RNA/DNA Complementarity.ACS Omega. 2019 Oct 9;4(17):17140-17147. doi: 10.1021/acsomega.9b01469. eCollection 2019 Oct 22. ACS Omega. 2019. PMID: 31656887 Free PMC article.
References
Methods-only References
-
- Huff J. The Airyscan detector from ZEISS: confocal imaging with improved signal-to-noise ratio and super-resolution. Nat Methods. 2015;12:i–ii.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
