

A Proteomics Survey of Junín Virus Interactions with Human Proteins Reveals Host Factors Required for Arenavirus Replication
- PMID: 29187543
- PMCID: PMC5790945
- DOI: 10.1128/JVI.01565-17
A Proteomics Survey of Junín Virus Interactions with Human Proteins Reveals Host Factors Required for Arenavirus Replication
Abstract
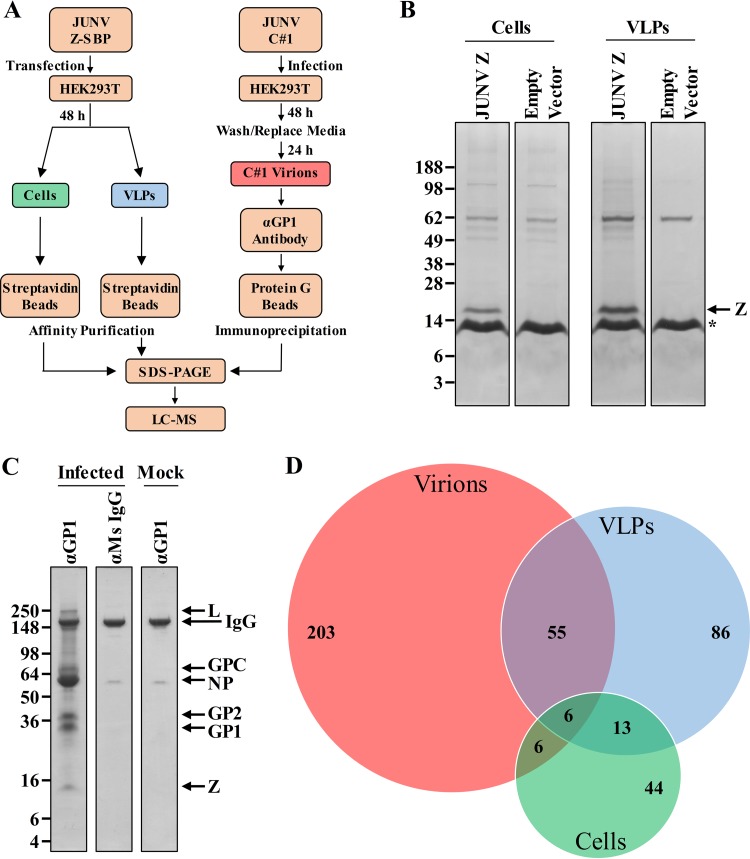
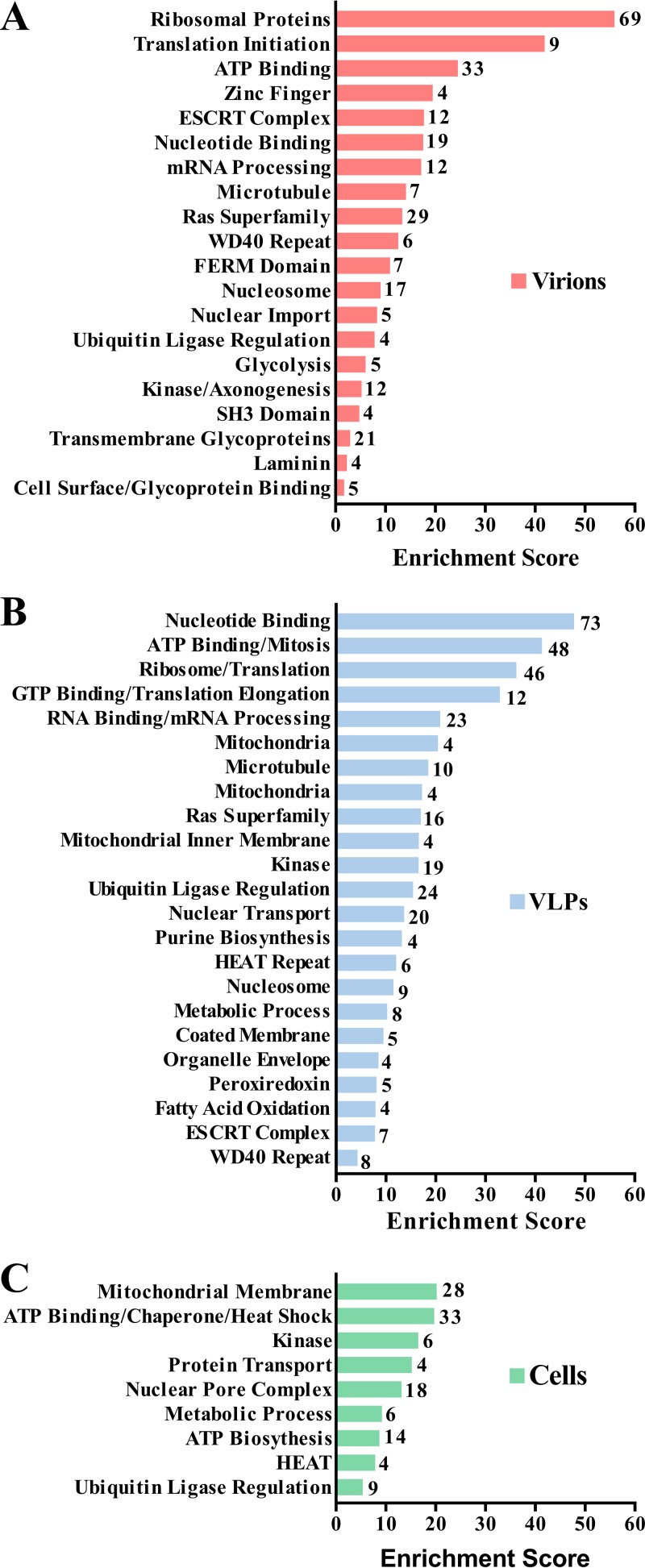
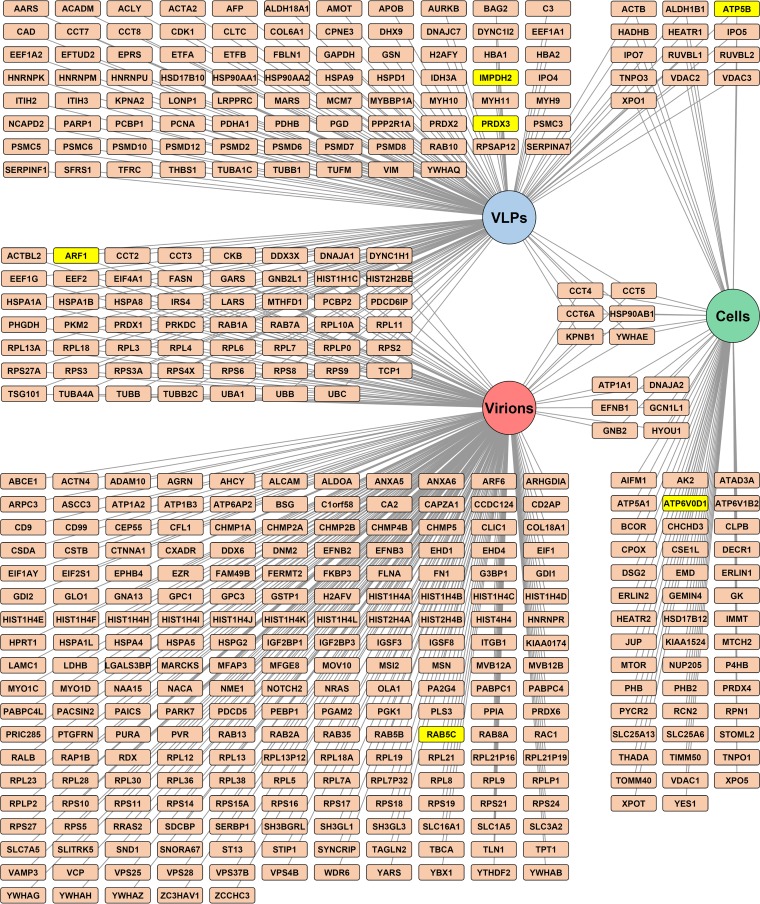
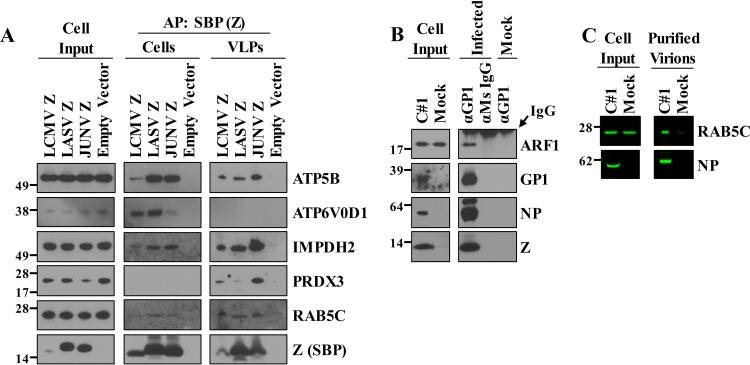
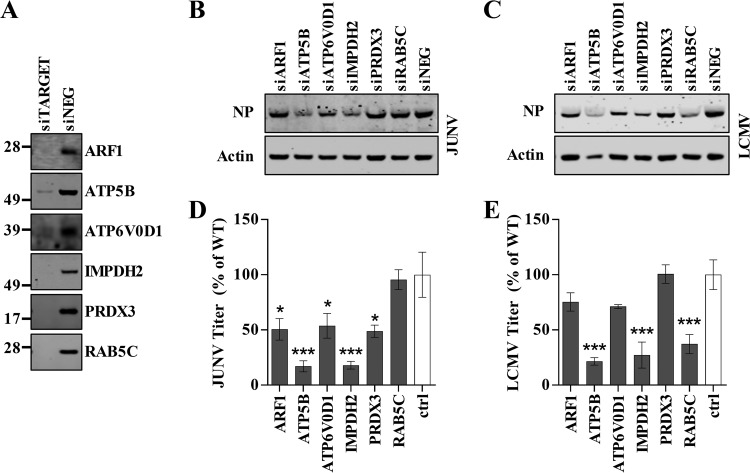
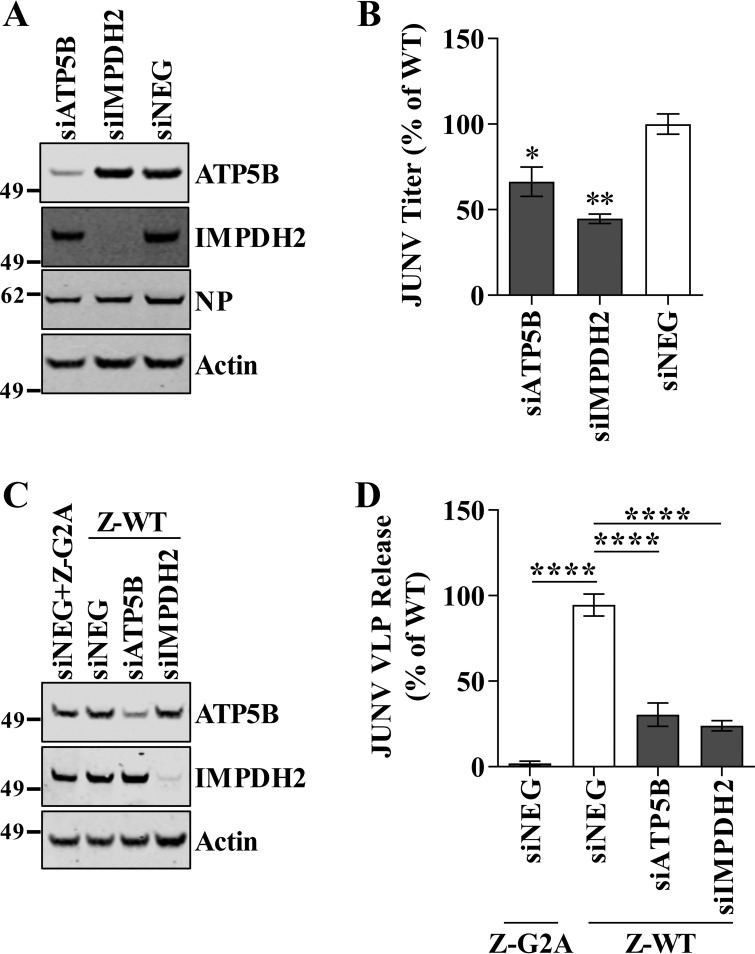
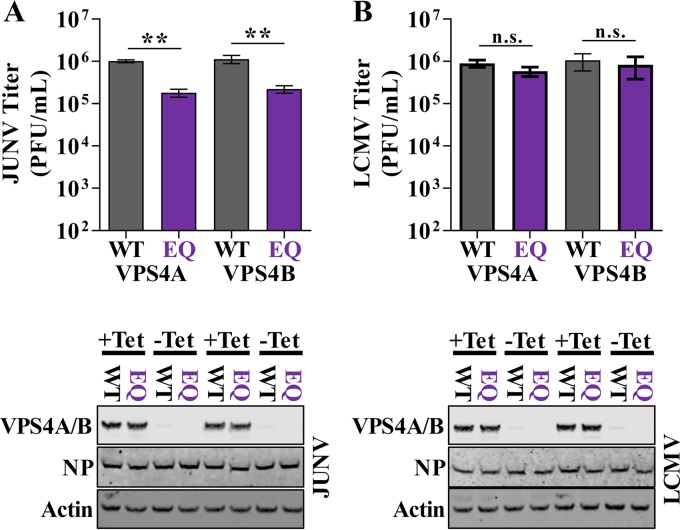
Arenaviruses are negative-strand, enveloped RNA viruses that cause significant human disease. In particular, Junín mammarenavirus (JUNV) is the etiologic agent of Argentine hemorrhagic fever. At present, little is known about the cellular proteins that the arenavirus matrix protein (Z) hijacks to accomplish its various functions, including driving the process of virus release. Furthermore, there is little knowledge regarding host proteins incorporated into arenavirus particles and their importance for virion function. To address these deficiencies, we used mass spectrometry to identify human proteins that (i) interact with the JUNV matrix protein inside cells or within virus-like particles (VLPs) and/or (ii) are incorporated into bona fide JUNV strain Candid#1 particles. Bioinformatics analyses revealed that multiple classes of human proteins were overrepresented in the data sets, including ribosomal proteins, Ras superfamily proteins, and endosomal sorting complex required for transport (ESCRT) proteins. Several of these proteins were required for the propagation of JUNV (ADP ribosylation factor 1 [ARF1], ATPase, H+ transporting, lysosomal 38-kDa, V0 subunit d1 [ATP6V0D1], and peroxiredoxin 3 [PRDX3]), lymphocytic choriomeningitis mammarenavirus (LCMV) (Rab5c), or both viruses (ATP synthase, H+ transporting, mitochondrial F1 complex, beta polypeptide [ATP5B] and IMP dehydrogenase 2 [IMPDH2]). Furthermore, we show that the release of infectious JUNV particles, but not LCMV particles, requires a functional ESCRT pathway and that ATP5B and IMPDH2 are required for JUNV budding. In summary, we have provided a large-scale map of host machinery that associates with JUNV and identified key human proteins required for its propagation. This data set provides a resource for the field to guide antiviral target discovery and to better understand the biology of the arenavirus matrix protein and the importance of host proteins for virion function.IMPORTANCE Arenaviruses are deadly human pathogens for which there are no U.S. Food and Drug Administration-approved vaccines and only limited treatment options. Little is known about the host proteins that are incorporated into arenavirus particles or that associate with its multifunctional matrix protein. Using Junín mammarenavirus (JUNV), the causative agent of Argentine hemorrhagic fever, as a model organism, we mapped the human proteins that are incorporated into JUNV particles or that associate with the JUNV matrix protein. Functional analysis revealed host machinery that is required for JUNV propagation, including the cellular ESCRT pathway. This study improves our understanding of critical arenavirus-host interactions and provides a data set that will guide future studies to better understand arenavirus pathogenesis and identify novel host proteins that can be therapeutically targeted.
Keywords: ESCRT; Junín virus; Rab5; VLP; Z; arenavirus; budding; interactome; lymphocytic choriomeningitis virus; mammarenavirus; matrix protein; matrix protein Z; protein-protein interactions; proteomics; virus particle.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
The Virus-Host Interplay in Junín Mammarenavirus Infection.Viruses. 2022 May 24;14(6):1134. doi: 10.3390/v14061134. Viruses. 2022. PMID: 35746604 Free PMC article. Review.
-
A Map of the Arenavirus Nucleoprotein-Host Protein Interactome Reveals that Junín Virus Selectively Impairs the Antiviral Activity of Double-Stranded RNA-Activated Protein Kinase (PKR).J Virol. 2017 Jul 12;91(15):e00763-17. doi: 10.1128/JVI.00763-17. Print 2017 Aug 1. J Virol. 2017. PMID: 28539447 Free PMC article.
-
A host-oriented inhibitor of Junin Argentine hemorrhagic fever virus egress.J Virol. 2014 May;88(9):4736-43. doi: 10.1128/JVI.03757-13. Epub 2014 Feb 12. J Virol. 2014. PMID: 24522922 Free PMC article.
-
Guinea Pig Transferrin Receptor 1 Mediates Cellular Entry of Junín Virus and Other Pathogenic New World Arenaviruses.J Virol. 2020 Jan 31;94(4):e01278-19. doi: 10.1128/JVI.01278-19. Print 2020 Jan 31. J Virol. 2020. PMID: 31748396 Free PMC article.
-
Arenavirus Quasispecies and Their Biological Implications.Curr Top Microbiol Immunol. 2016;392:231-76. doi: 10.1007/82_2015_468. Curr Top Microbiol Immunol. 2016. PMID: 26472215 Free PMC article. Review.
Cited by
-
Lifecycle modelling systems support inosine monophosphate dehydrogenase (IMPDH) as a pro-viral factor and antiviral target for New World arenaviruses.Antiviral Res. 2018 Sep;157:140-150. doi: 10.1016/j.antiviral.2018.07.009. Epub 2018 Jul 19. Antiviral Res. 2018. PMID: 30031760 Free PMC article.
-
The Virus-Host Interplay in Junín Mammarenavirus Infection.Viruses. 2022 May 24;14(6):1134. doi: 10.3390/v14061134. Viruses. 2022. PMID: 35746604 Free PMC article. Review.
-
Proteomic Profiling of Purified Rabies Virus Particles.Virol Sin. 2020 Apr;35(2):143-155. doi: 10.1007/s12250-019-00157-6. Epub 2019 Aug 19. Virol Sin. 2020. PMID: 31429011 Free PMC article.
-
SARS-CoV-2 Nsp14 activates NF-κB signaling and induces IL-8 upregulation.bioRxiv [Preprint]. 2021 May 26:2021.05.26.445787. doi: 10.1101/2021.05.26.445787. bioRxiv. 2021. PMID: 34075374 Free PMC article. Preprint.
-
Virus⁻Host Interactions Involved in Lassa Virus Entry and Genome Replication.Pathogens. 2019 Jan 29;8(1):17. doi: 10.3390/pathogens8010017. Pathogens. 2019. PMID: 30699976 Free PMC article. Review.
References
-
- Salazar-Bravo J, Ruedas LA, Yates TL. 2002. Mammalian reservoirs of arenaviruses. Curr Top Microbiol Immunol 262:25–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
