

Direction of leukocyte polarization and migration by the phosphoinositide-transfer protein TIPE2
- PMID: 29058702
- PMCID: PMC5690821
- DOI: 10.1038/ni.3866
Direction of leukocyte polarization and migration by the phosphoinositide-transfer protein TIPE2
Abstract
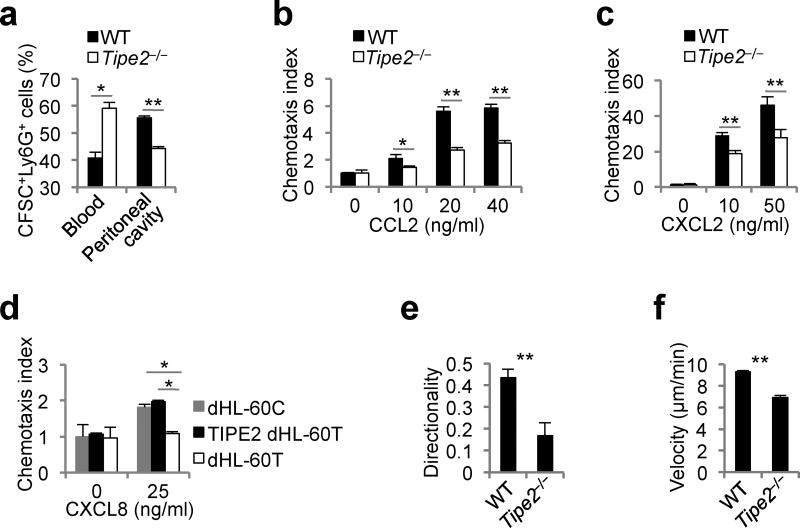
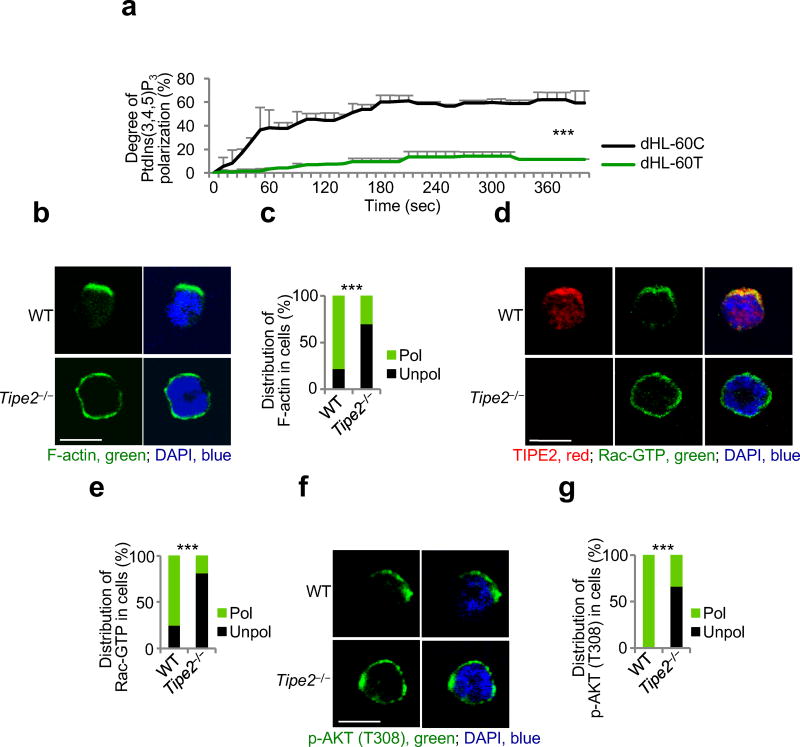
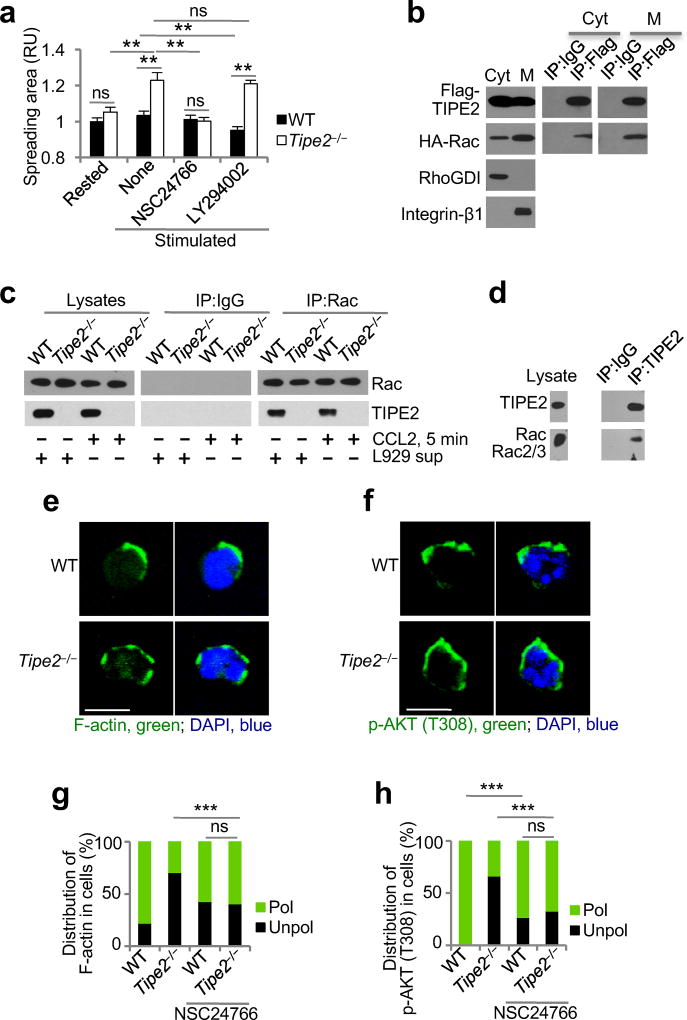
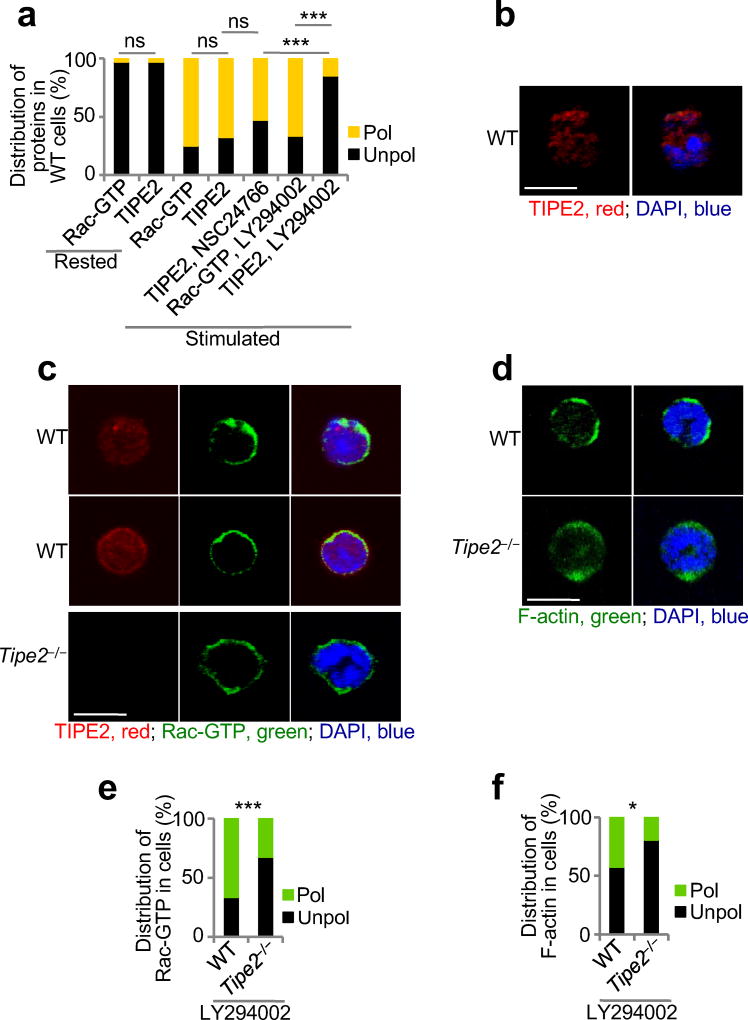
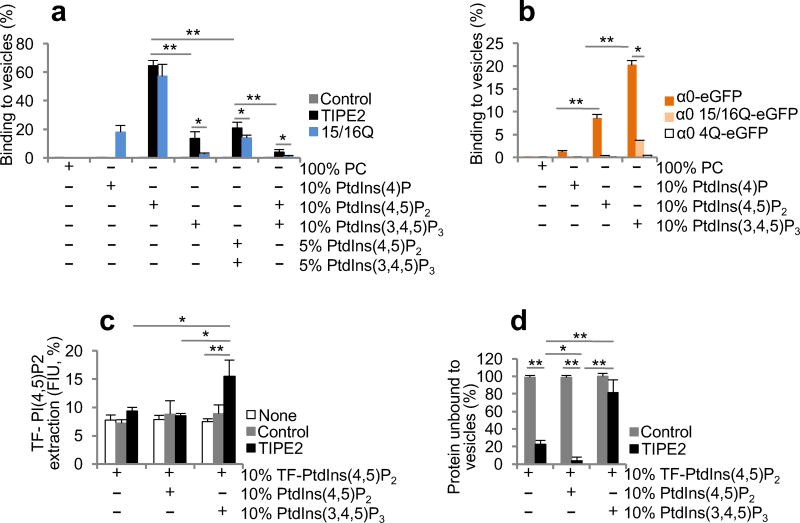
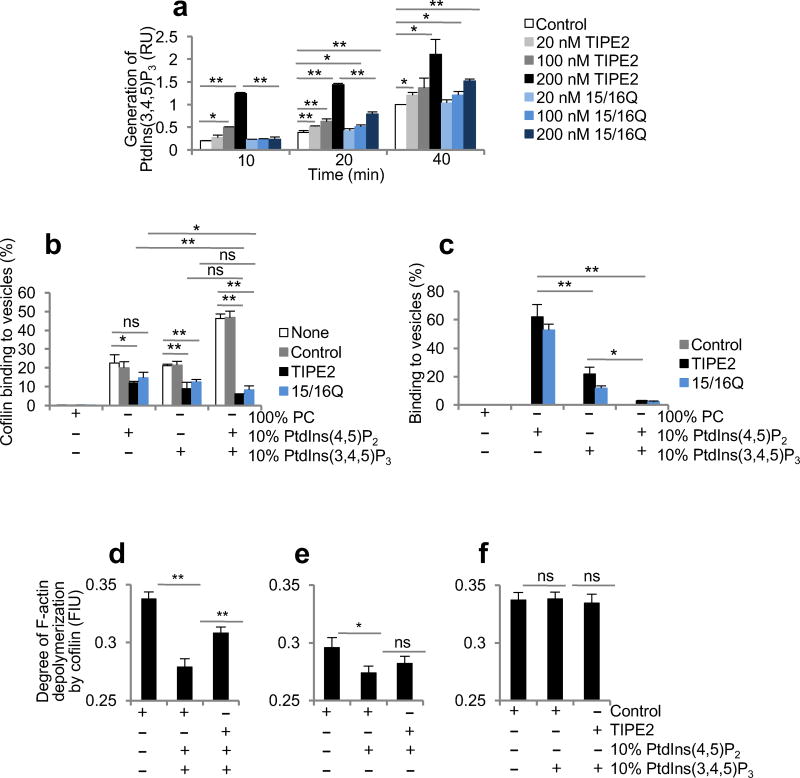
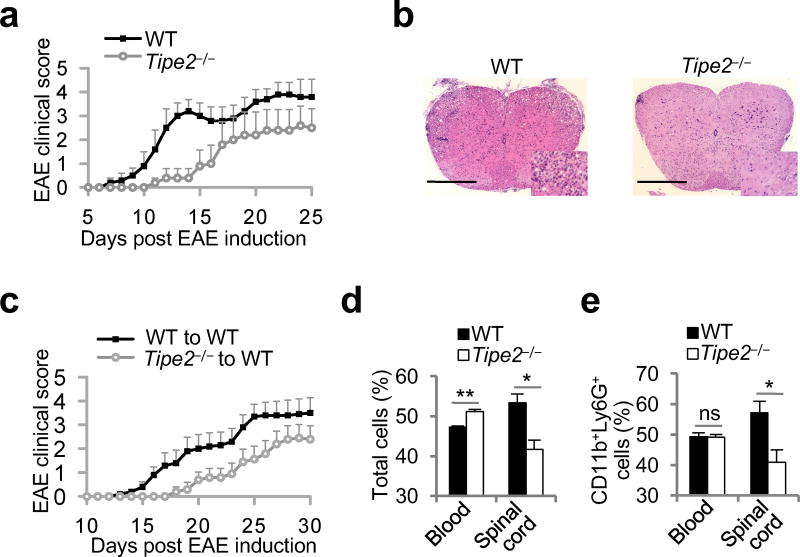
The polarization of leukocytes toward chemoattractants is essential for the directed migration (chemotaxis) of leukocytes. How leukocytes acquire polarity after encountering chemical gradients is not well understood. We found here that leukocyte polarity was generated by TIPE2 (TNFAIP8L2), a transfer protein for phosphoinositide second messengers. TIPE2 functioned as a local enhancer of phosphoinositide-dependent signaling and cytoskeleton remodeling, which promoted leading-edge formation. Conversely, TIPE2 acted as an inhibitor of the GTPase Rac, which promoted trailing-edge polarization. Consequently, TIPE2-deficient leukocytes were defective in polarization and chemotaxis, and TIPE2-deficient mice were resistant to leukocyte-mediated neural inflammation. Thus, the leukocyte polarizer is a dual-role phosphoinositide-transfer protein and represents a potential therapeutic target for the treatment of inflammatory diseases.
Conflict of interest statement
The authors have no financial conflict of interest with this work.
Figures
Similar articles
-
TIPE2 controls innate immunity to RNA by targeting the phosphatidylinositol 3-kinase-Rac pathway.J Immunol. 2012 Sep 15;189(6):2768-73. doi: 10.4049/jimmunol.1103477. Epub 2012 Aug 17. J Immunol. 2012. PMID: 22904303 Free PMC article.
-
TIPE2 specifies the functional polarization of myeloid-derived suppressor cells during tumorigenesis.J Exp Med. 2020 Feb 3;217(2):e20182005. doi: 10.1084/jem.20182005. J Exp Med. 2020. PMID: 31662347 Free PMC article.
-
Egress of murine regulatory T cells from the thymus requires TIPE2.Biochem Biophys Res Commun. 2018 Jun 2;500(2):376-383. doi: 10.1016/j.bbrc.2018.04.082. Epub 2018 Apr 14. Biochem Biophys Res Commun. 2018. PMID: 29654762
-
Do phosphoinositide 3-kinases direct lymphocyte navigation?Trends Immunol. 2004 Feb;25(2):67-74. doi: 10.1016/j.it.2003.12.003. Trends Immunol. 2004. PMID: 15102365 Review.
-
Lipids on the move: phosphoinositide 3-kinases in leukocyte function.Immunol Today. 2000 Jun;21(6):260-4. doi: 10.1016/s0167-5699(00)01649-2. Immunol Today. 2000. PMID: 10939787 Review. No abstract available.
Cited by
-
Decoupling tumor cell metastasis from growth by cellular pilot protein TNFAIP8.Oncogene. 2021 Nov;40(46):6456-6468. doi: 10.1038/s41388-021-02035-6. Epub 2021 Oct 4. Oncogene. 2021. PMID: 34608264 Free PMC article.
-
TIPE2 is a checkpoint of natural killer cell maturation and antitumor immunity.Sci Adv. 2021 Sep 17;7(38):eabi6515. doi: 10.1126/sciadv.abi6515. Epub 2021 Sep 15. Sci Adv. 2021. PMID: 34524845 Free PMC article.
-
Tnfaip8 and Tipe2 Gene Deletion Ameliorates Immediate Proteoglycan Loss and Inflammatory Responses in the Injured Mouse Intervertebral Disc.Am J Phys Med Rehabil. 2024 Oct 1;103(10):918-924. doi: 10.1097/PHM.0000000000002488. Epub 2024 Apr 8. Am J Phys Med Rehabil. 2024. PMID: 38630557
-
Streamlining Computational Fragment-Based Drug Discovery through Evolutionary Optimization Informed by Ligand-Based Virtual Prescreening.J Chem Inf Model. 2024 May 13;64(9):3826-3840. doi: 10.1021/acs.jcim.4c00234. Epub 2024 May 2. J Chem Inf Model. 2024. PMID: 38696451 Free PMC article.
-
TIPE1 Suppresses Growth and Metastasis of Ovarian Cancer.J Oncol. 2021 Jun 3;2021:5538911. doi: 10.1155/2021/5538911. eCollection 2021. J Oncol. 2021. PMID: 34188681 Free PMC article.
References
-
- Merlot S, Firtel RA. Leading the way: Directional sensing through phosphatidylinositol 3-kinase and other signaling pathways. J. Cell Sci. 2003;116:3471–3478. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
