

Architectural alterations of the fission yeast genome during the cell cycle
- PMID: 28991264
- PMCID: PMC5724045
- DOI: 10.1038/nsmb.3482
Architectural alterations of the fission yeast genome during the cell cycle
Abstract
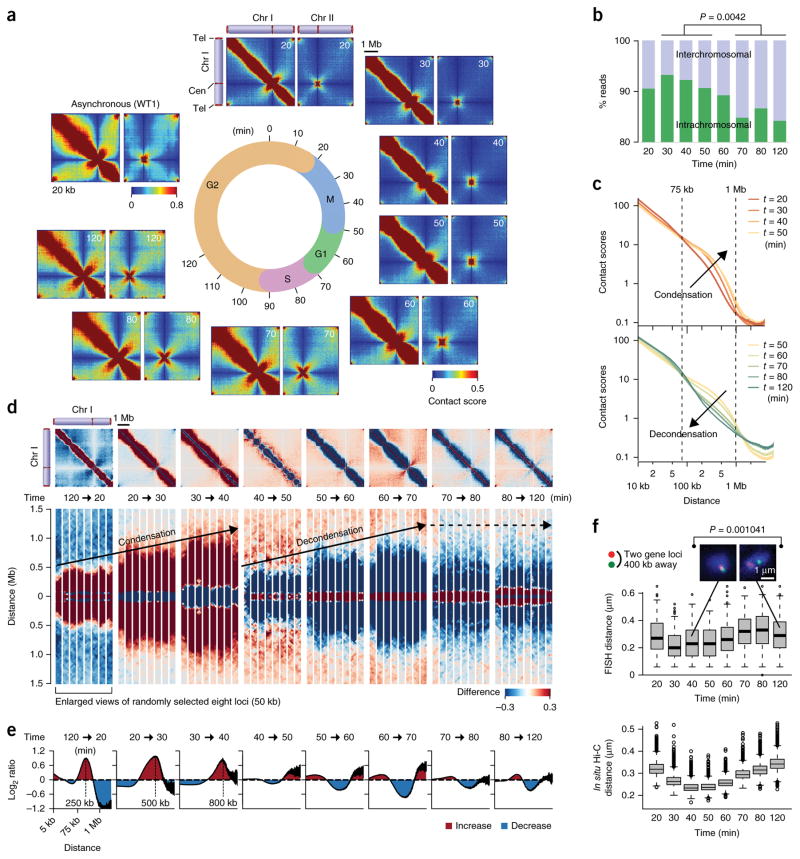
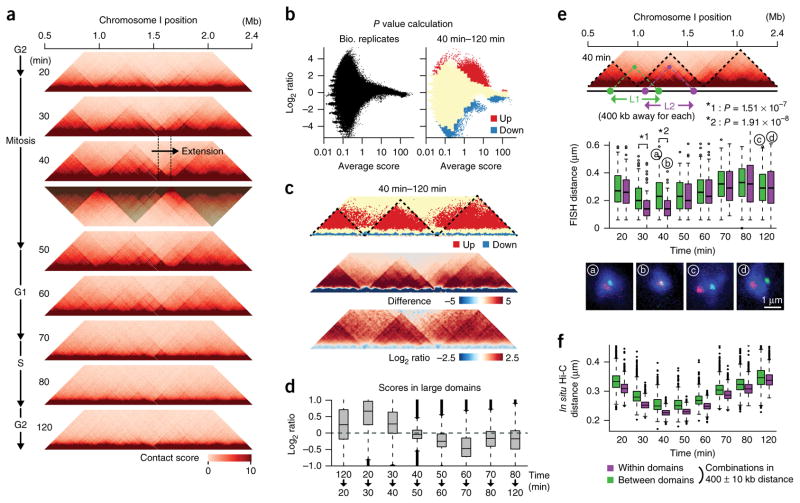
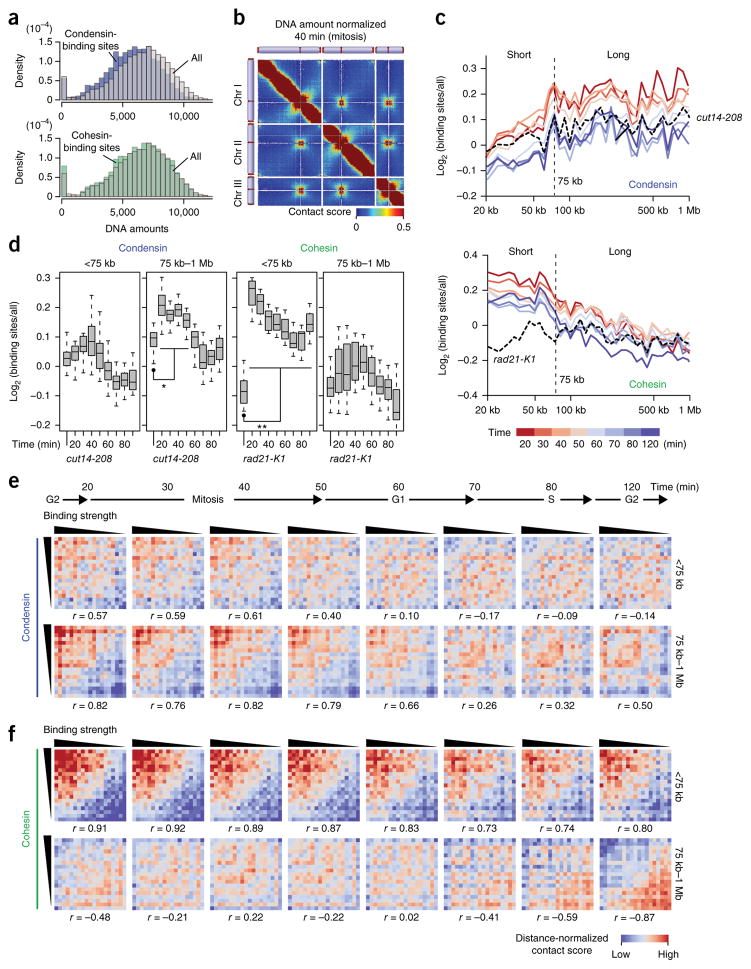
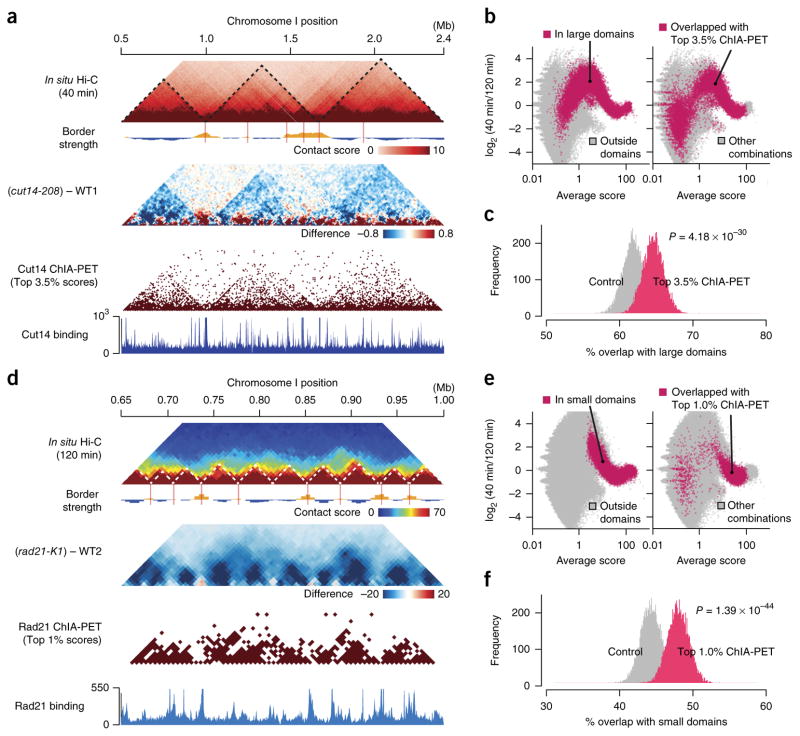
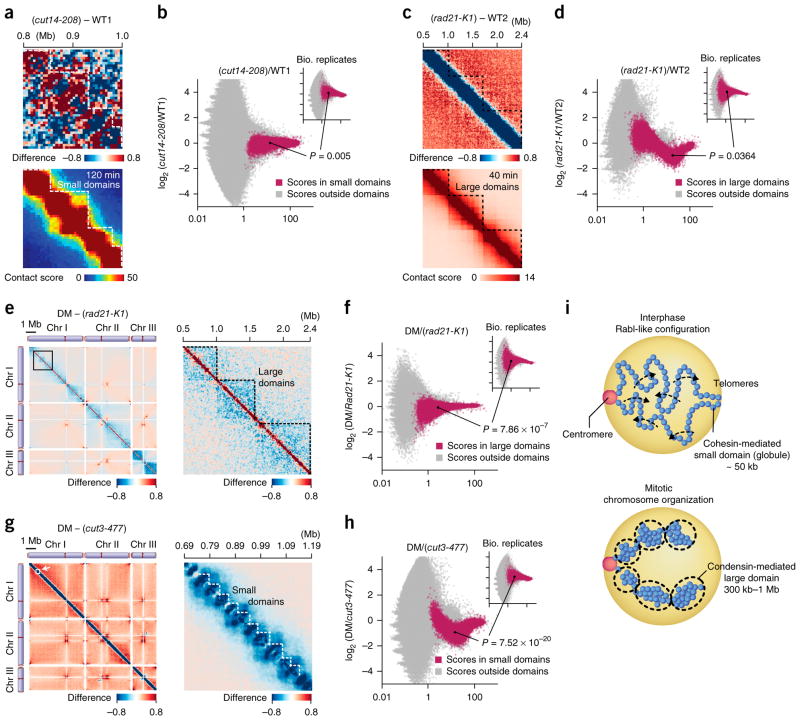
Eukaryotic genomes are highly ordered through various mechanisms, including topologically associating domain (TAD) organization. We employed an in situ Hi-C approach to follow the 3D organization of the fission yeast genome during the cell cycle. We demonstrate that during mitosis, large domains of 300 kb-1 Mb are formed by condensin. This mitotic domain organization does not suddenly dissolve, but gradually diminishes until the next mitosis. By contrast, small domains of 30-40 kb that are formed by cohesin are relatively stable across the cell cycle. Condensin and cohesin mediate long- and short-range contacts, respectively, by bridging their binding sites, thereby forming the large and small domains. These domains are inversely regulated during the cell cycle but assemble independently. Our study describes the chromosomal oscillation between the formation and decay phases of the large and small domains, and we predict that the condensin-mediated domains serve as chromosomal compaction units.
Conflict of interest statement
The authors declare no competing financial interests.
Figures


Similar articles
-
Potential roles of condensin in genome organization and beyond in fission yeast.J Microbiol. 2021 May;59(5):449-459. doi: 10.1007/s12275-021-1039-2. Epub 2021 Apr 20. J Microbiol. 2021. PMID: 33877578 Review.
-
Transcription factors mediate condensin recruitment and global chromosomal organization in fission yeast.Nat Genet. 2016 Oct;48(10):1242-52. doi: 10.1038/ng.3647. Epub 2016 Aug 22. Nat Genet. 2016. PMID: 27548313 Free PMC article.
-
The Yeast Genomes in Three Dimensions: Mechanisms and Functions.Annu Rev Genet. 2017 Nov 27;51:23-44. doi: 10.1146/annurev-genet-120116-023438. Epub 2017 Aug 30. Annu Rev Genet. 2017. PMID: 28853923 Review.
-
Condensin-mediated chromosome organization in fission yeast.Curr Genet. 2016 Nov;62(4):739-743. doi: 10.1007/s00294-016-0601-7. Epub 2016 Apr 9. Curr Genet. 2016. PMID: 27061734 Free PMC article. Review.
-
Nucleosome eviction in mitosis assists condensin loading and chromosome condensation.EMBO J. 2016 Jul 15;35(14):1565-81. doi: 10.15252/embj.201592849. Epub 2016 Jun 6. EMBO J. 2016. PMID: 27266525 Free PMC article.
Cited by
-
Condensin-Dependent Chromatin Compaction Represses Transcription Globally during Quiescence.Mol Cell. 2019 Feb 7;73(3):533-546.e4. doi: 10.1016/j.molcel.2018.11.020. Epub 2018 Dec 27. Mol Cell. 2019. PMID: 30595435 Free PMC article.
-
A mini-review of the role of condensin in human nervous system diseases.Front Mol Neurosci. 2022 Aug 4;15:889796. doi: 10.3389/fnmol.2022.889796. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35992200 Free PMC article. Review.
-
Histone deacetylation and cytosine methylation compartmentalize heterochromatic regions in the genome organization of Neurospora crassa.Proc Natl Acad Sci U S A. 2023 Nov 21;120(47):e2311249120. doi: 10.1073/pnas.2311249120. Epub 2023 Nov 14. Proc Natl Acad Sci U S A. 2023. PMID: 37963248 Free PMC article.
-
A compendium of chromatin contact maps reflecting regulation by chromatin remodelers in budding yeast.Nat Commun. 2021 Nov 4;12(1):6380. doi: 10.1038/s41467-021-26629-6. Nat Commun. 2021. PMID: 34737268 Free PMC article.
-
Potential roles of condensin in genome organization and beyond in fission yeast.J Microbiol. 2021 May;59(5):449-459. doi: 10.1007/s12275-021-1039-2. Epub 2021 Apr 20. J Microbiol. 2021. PMID: 33877578 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
