

Nesprin-1α-Dependent Microtubule Nucleation from the Nuclear Envelope via Akap450 Is Necessary for Nuclear Positioning in Muscle Cells
- PMID: 28966089
- PMCID: PMC5640514
- DOI: 10.1016/j.cub.2017.08.031
Nesprin-1α-Dependent Microtubule Nucleation from the Nuclear Envelope via Akap450 Is Necessary for Nuclear Positioning in Muscle Cells
Abstract
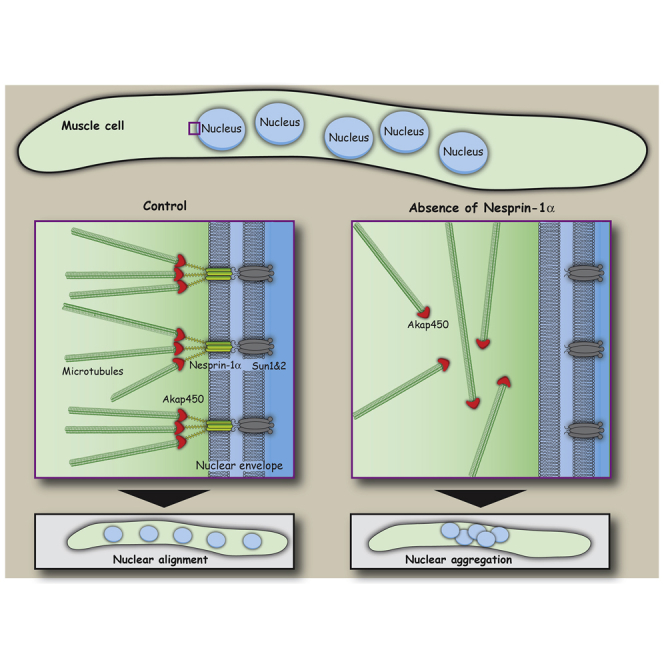
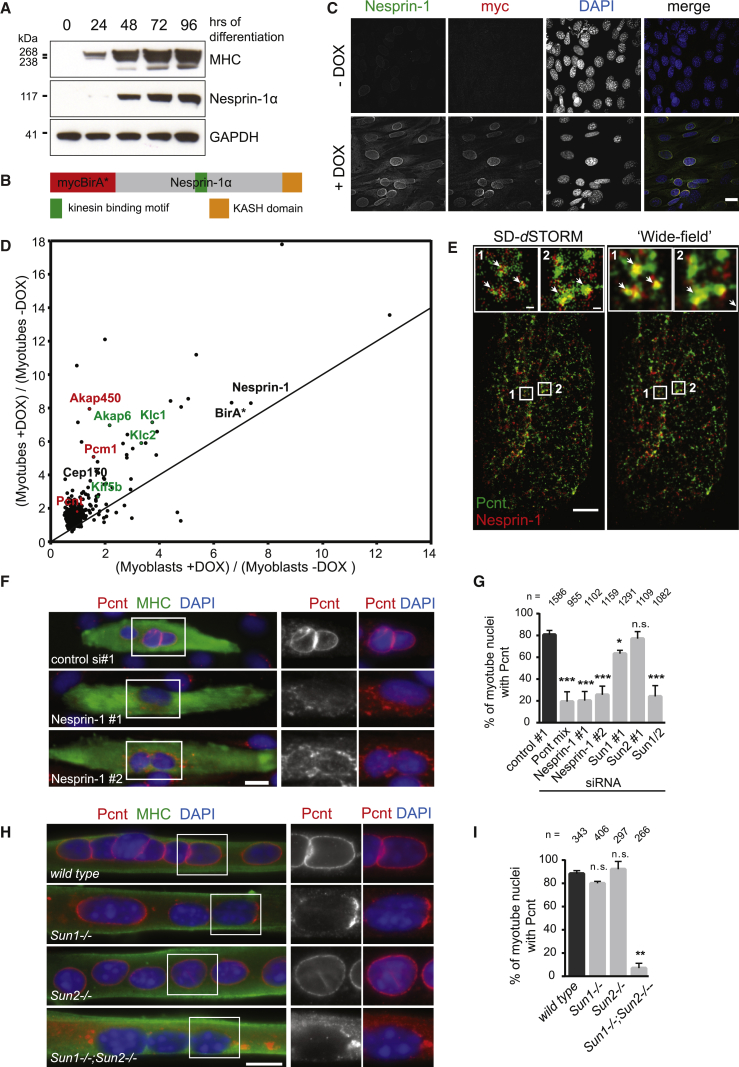
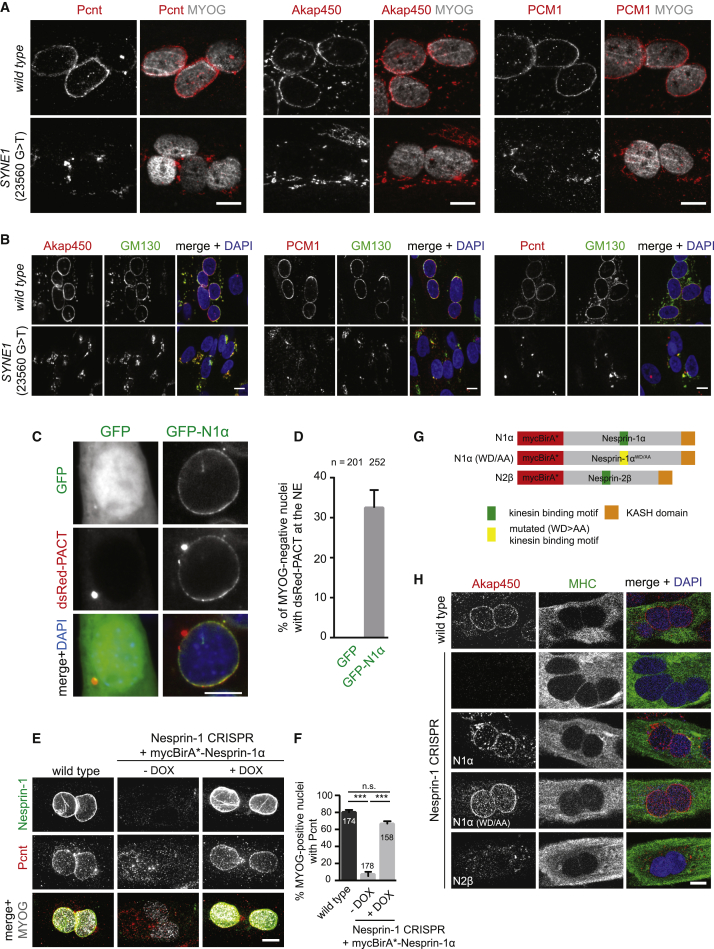
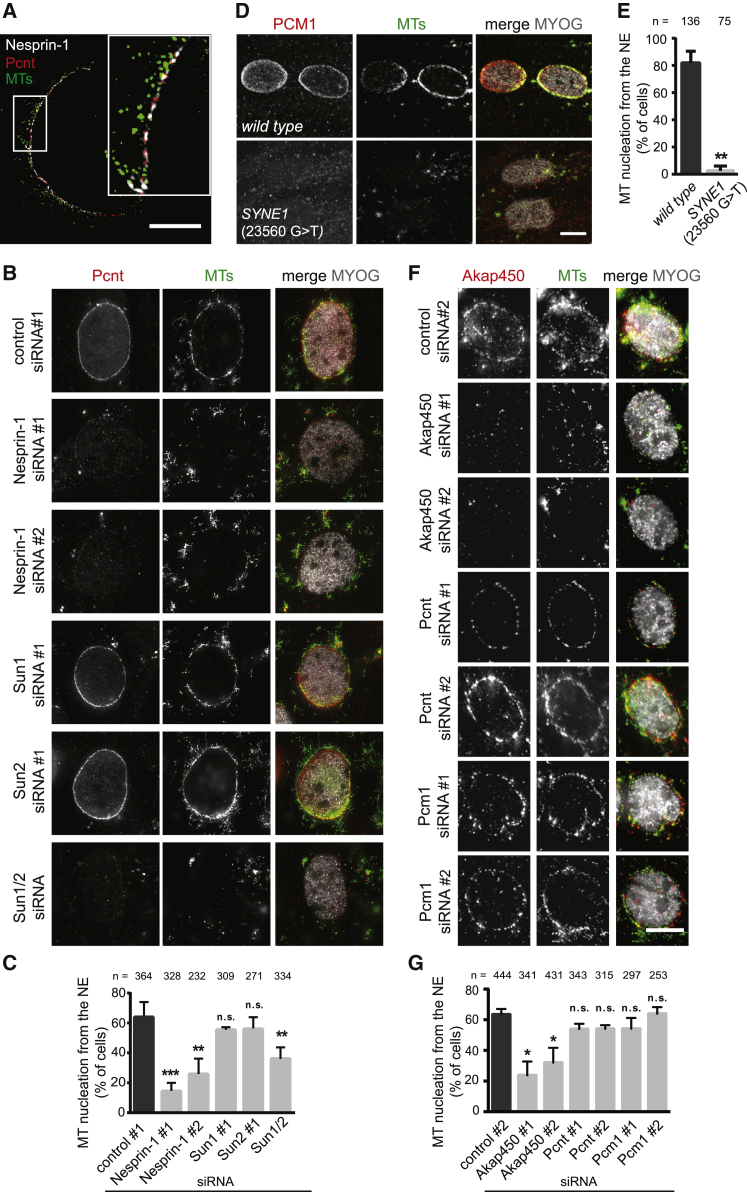
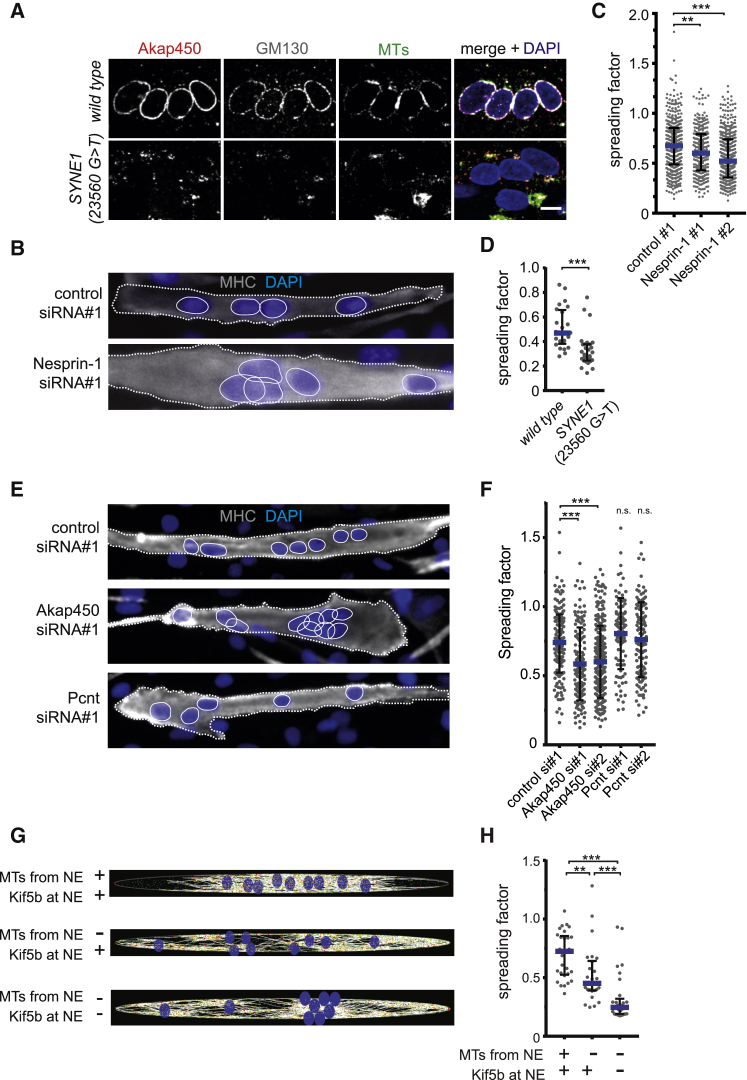
The nucleus is the main microtubule-organizing center (MTOC) in muscle cells due to the accumulation of centrosomal proteins and microtubule (MT) nucleation activity at the nuclear envelope (NE) [1-4]. The relocalization of centrosomal proteins, including Pericentrin, Pcm1, and γ-tubulin, depends on Nesprin-1, an outer nuclear membrane (ONM) protein that connects the nucleus to the cytoskeleton via its N-terminal region [5-7]. Nesprins are also involved in the recruitment of kinesin to the NE and play a role in nuclear positioning in skeletal muscle cells [8-12]. However, a function for MT nucleation from the NE in nuclear positioning has not been established. Using the proximity-dependent biotin identification (BioID) method [13, 14], we found several centrosomal proteins, including Akap450, Pcm1, and Pericentrin, whose association with Nesprin-1α is increased in differentiated myotubes. We show that Nesprin-1α recruits Akap450 to the NE independently of kinesin and that Akap450, but not other centrosomal proteins, is required for MT nucleation from the NE. Furthermore, we demonstrate that this mechanism is disrupted in congenital muscular dystrophy patient myotubes carrying a nonsense mutation within the SYNE1 gene (23560 G>T) encoding Nesprin-1 [15, 16]. Finally, using computer simulation and cell culture systems, we provide evidence for a role of MT nucleation from the NE on nuclear spreading in myotubes. Our data thus reveal a novel function for Nesprin-1α/Nesprin-1 in nuclear positioning through recruitment of Akap450-mediated MT nucleation activity to the NE.
Keywords: Akap450; Nesprin-1; Nesprin-1α; centrosome; cytosim computer simulation; microtubules; non-centrosomal MTOC; nuclear envelope; nuclear positioning; skeletal muscle.
Copyright © 2017 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Muscle Development: Nucleating Microtubules at the Nuclear Envelope.Curr Biol. 2017 Oct 9;27(19):R1071-R1073. doi: 10.1016/j.cub.2017.08.030. Curr Biol. 2017. PMID: 29017044 Free PMC article.
Similar articles
-
AKAP6 orchestrates the nuclear envelope microtubule-organizing center by linking golgi and nucleus via AKAP9.Elife. 2020 Dec 9;9:e61669. doi: 10.7554/eLife.61669. Elife. 2020. PMID: 33295871 Free PMC article.
-
Novel nesprin-1 mutations associated with dilated cardiomyopathy cause nuclear envelope disruption and defects in myogenesis.Hum Mol Genet. 2017 Jun 15;26(12):2258-2276. doi: 10.1093/hmg/ddx116. Hum Mol Genet. 2017. PMID: 28398466 Free PMC article.
-
Microtubule nucleation at the cis-side of the Golgi apparatus requires AKAP450 and GM130.EMBO J. 2009 Apr 22;28(8):1016-28. doi: 10.1038/emboj.2009.47. Epub 2009 Feb 26. EMBO J. 2009. PMID: 19242490 Free PMC article.
-
Nesprin-1: novel regulator of striated muscle nuclear positioning and mechanotransduction.Biochem Soc Trans. 2023 Jun 28;51(3):1331-1345. doi: 10.1042/BST20221541. Biochem Soc Trans. 2023. PMID: 37171063 Free PMC article. Review.
-
Regulation of microtubule nucleation mediated by γ-tubulin complexes.Protoplasma. 2017 May;254(3):1187-1199. doi: 10.1007/s00709-016-1070-z. Epub 2017 Jan 10. Protoplasma. 2017. PMID: 28074286 Review.
Cited by
-
The nuclear envelope: LINCing tissue mechanics to genome regulation in cardiac and skeletal muscle.Biol Lett. 2020 Jul;16(7):20200302. doi: 10.1098/rsbl.2020.0302. Epub 2020 Jul 8. Biol Lett. 2020. PMID: 32634376 Free PMC article.
-
The Pathogenesis and Therapies of Striated Muscle Laminopathies.Front Physiol. 2018 Oct 30;9:1533. doi: 10.3389/fphys.2018.01533. eCollection 2018. Front Physiol. 2018. PMID: 30425656 Free PMC article. Review.
-
Atrial Structural Remodeling Gene Variants in Patients with Atrial Fibrillation.Biomed Res Int. 2018 Sep 10;2018:4862480. doi: 10.1155/2018/4862480. eCollection 2018. Biomed Res Int. 2018. PMID: 30276209 Free PMC article.
-
Nesprin-1 LINC complexes recruit microtubule cytoskeleton proteins and drive pathology in Lmna-mutant striated muscle.Hum Mol Genet. 2023 Jan 6;32(2):177-191. doi: 10.1093/hmg/ddac179. Hum Mol Genet. 2023. PMID: 35925868 Free PMC article.
-
Unraveling the mysteries of centriolar satellites: time to rewrite the textbooks about the centrosome/cilium complex.Mol Biol Cell. 2020 Apr 15;31(9):866-872. doi: 10.1091/mbc.E19-07-0402. Mol Biol Cell. 2020. PMID: 32286929 Free PMC article. Review.
References
-
- Bugnard E., Zaal K.J.M., Ralston E. Reorganization of microtubule nucleation during muscle differentiation. Cell Motil. Cytoskeleton. 2005;60:1–13. - PubMed
-
- Espigat-Georger A., Dyachuk V., Chemin C., Emorine L., Merdes A. Nuclear alignment in myotubes requires centrosome proteins recruited by nesprin-1. J. Cell Sci. 2016;129:4227–4237. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
