

A general mechanism of ribosome dimerization revealed by single-particle cryo-electron microscopy
- PMID: 28959045
- PMCID: PMC5620043
- DOI: 10.1038/s41467-017-00718-x
A general mechanism of ribosome dimerization revealed by single-particle cryo-electron microscopy
Abstract
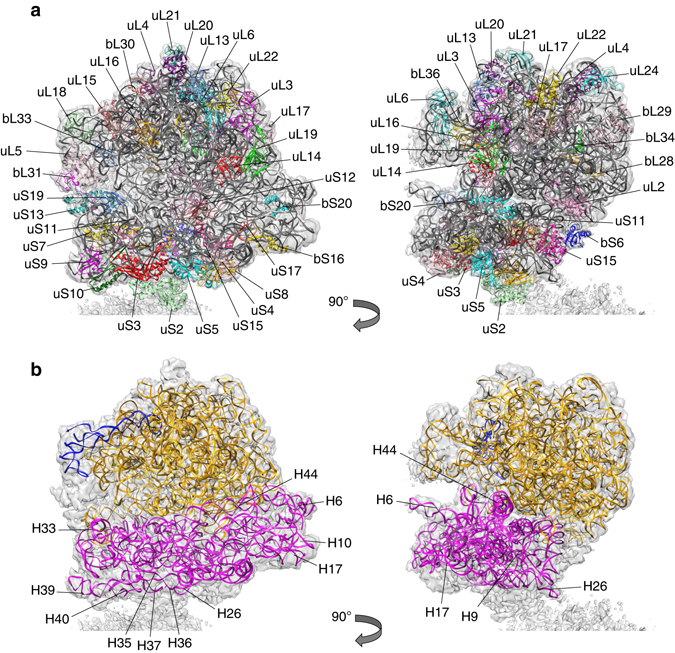
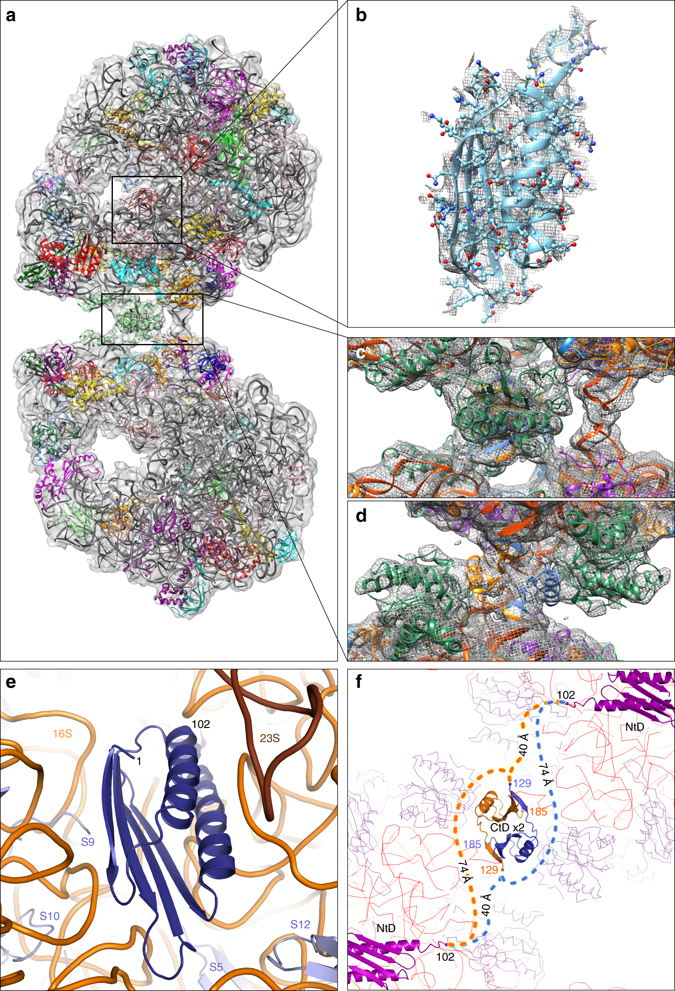
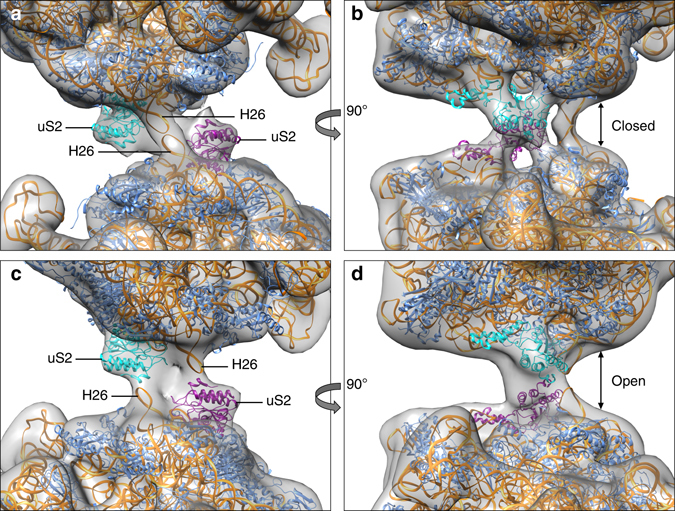
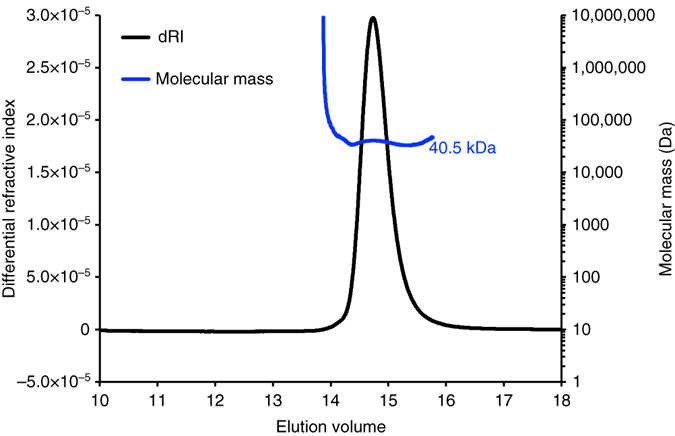
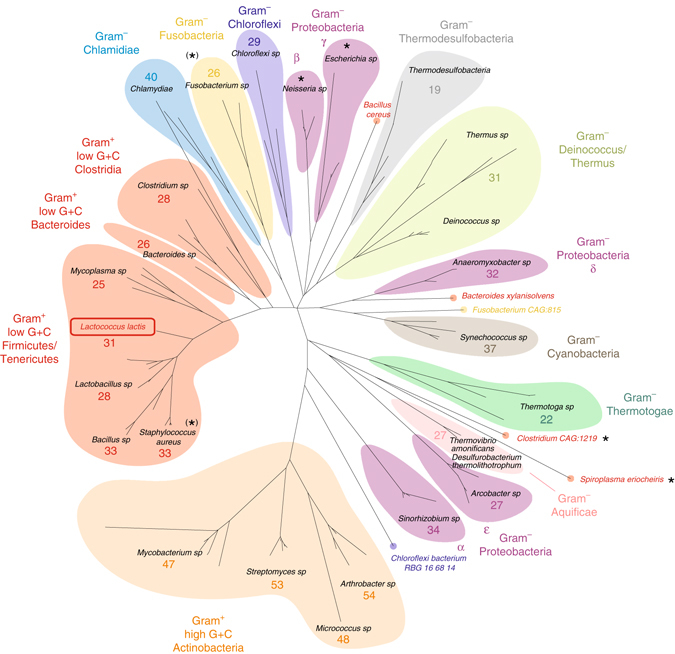
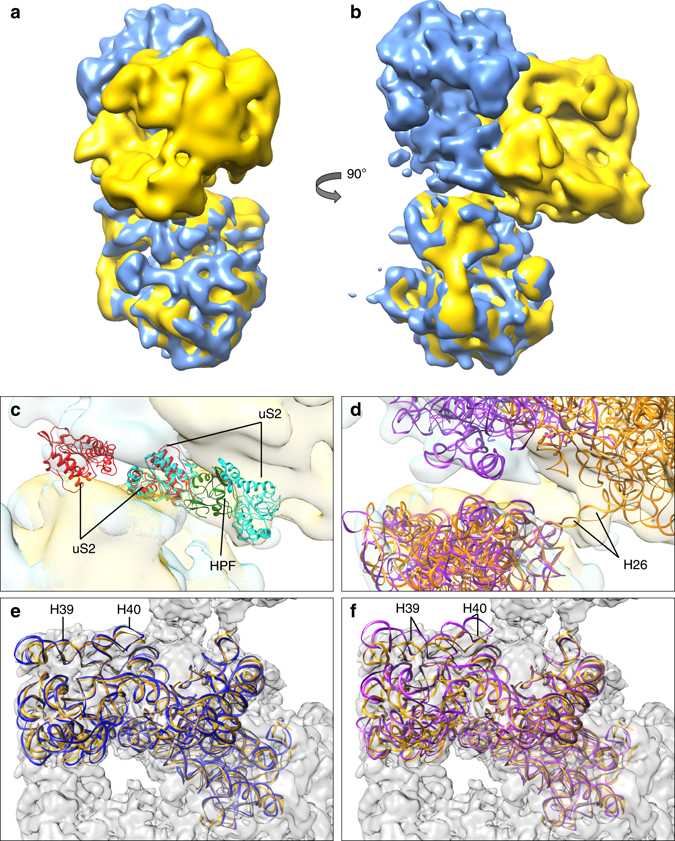
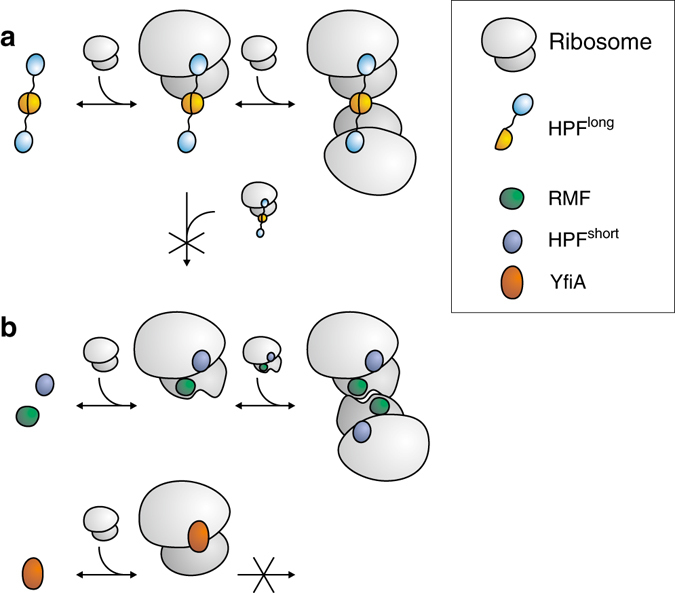
Bacteria downregulate their ribosomal activity through dimerization of 70S ribosomes, yielding inactive 100S complexes. In Escherichia coli, dimerization is mediated by the hibernation promotion factor (HPF) and ribosome modulation factor. Here we report the cryo-electron microscopy study on 100S ribosomes from Lactococcus lactis and a dimerization mechanism involving a single protein: HPFlong. The N-terminal domain of HPFlong binds at the same site as HPF in Escherichia coli 100S ribosomes. Contrary to ribosome modulation factor, the C-terminal domain of HPFlong binds exactly at the dimer interface. Furthermore, ribosomes from Lactococcus lactis do not undergo conformational changes in the 30S head domains upon binding of HPFlong, and the Shine-Dalgarno sequence and mRNA entrance tunnel remain accessible. Ribosome activity is blocked by HPFlong due to the inhibition of mRNA recognition by the platform binding center. Phylogenetic analysis of HPF proteins suggests that HPFlong-mediated dimerization is a widespread mechanism of ribosome hibernation in bacteria.When bacteria enter the stationary growth phase, protein translation is suppressed via the dimerization of 70S ribosomes into inactive complexes. Here the authors provide a structural basis for how the dual domain hibernation promotion factor promotes ribosome dimerization and hibernation in bacteria.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Stress response as implemented by hibernating ribosomes: a structural overview.FEBS J. 2019 Sep;286(18):3558-3565. doi: 10.1111/febs.14968. Epub 2019 Jul 15. FEBS J. 2019. PMID: 31230411 Free PMC article. Review.
-
Lactococcus lactis YfiA is necessary and sufficient for ribosome dimerization.Mol Microbiol. 2014 Jan;91(2):394-407. doi: 10.1111/mmi.12468. Epub 2013 Dec 13. Mol Microbiol. 2014. PMID: 24279750
-
Structure of a hibernating 100S ribosome reveals an inactive conformation of the ribosomal protein S1.Nat Microbiol. 2018 Oct;3(10):1115-1121. doi: 10.1038/s41564-018-0237-0. Epub 2018 Sep 3. Nat Microbiol. 2018. PMID: 30177741
-
Structure of the Bacillus subtilis hibernating 100S ribosome reveals the basis for 70S dimerization.EMBO J. 2017 Jul 14;36(14):2061-2072. doi: 10.15252/embj.201696189. Epub 2017 May 3. EMBO J. 2017. PMID: 28468753 Free PMC article.
-
Survival of the drowsiest: the hibernating 100S ribosome in bacterial stress management.Curr Genet. 2018 Aug;64(4):753-760. doi: 10.1007/s00294-017-0796-2. Epub 2017 Dec 14. Curr Genet. 2018. PMID: 29243175 Free PMC article. Review.
Cited by
-
Cryo-EM structure of the hibernating Thermus thermophilus 100S ribosome reveals a protein-mediated dimerization mechanism.Nat Commun. 2018 Oct 9;9(1):4179. doi: 10.1038/s41467-018-06724-x. Nat Commun. 2018. PMID: 30301898 Free PMC article.
-
Phage-Related Ribosomal Proteases (Prps): Discovery, Bioinformatics, and Structural Analysis.Antibiotics (Basel). 2022 Aug 16;11(8):1109. doi: 10.3390/antibiotics11081109. Antibiotics (Basel). 2022. PMID: 36009978 Free PMC article.
-
Stress response as implemented by hibernating ribosomes: a structural overview.FEBS J. 2019 Sep;286(18):3558-3565. doi: 10.1111/febs.14968. Epub 2019 Jul 15. FEBS J. 2019. PMID: 31230411 Free PMC article. Review.
-
Ribosomal Hibernation-Associated Factors in Escherichia coli.Microorganisms. 2021 Dec 24;10(1):33. doi: 10.3390/microorganisms10010033. Microorganisms. 2021. PMID: 35056482 Free PMC article. Review.
-
Sleeping ribosomes: Bacterial signaling triggers RaiA mediated persistence to aminoglycosides.iScience. 2021 Sep 14;24(10):103128. doi: 10.1016/j.isci.2021.103128. eCollection 2021 Oct 22. iScience. 2021. PMID: 34611612 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
