

Mettl3-/Mettl14-mediated mRNA N6-methyladenosine modulates murine spermatogenesis
- PMID: 28914256
- PMCID: PMC5630681
- DOI: 10.1038/cr.2017.117
Mettl3-/Mettl14-mediated mRNA N6-methyladenosine modulates murine spermatogenesis
Abstract
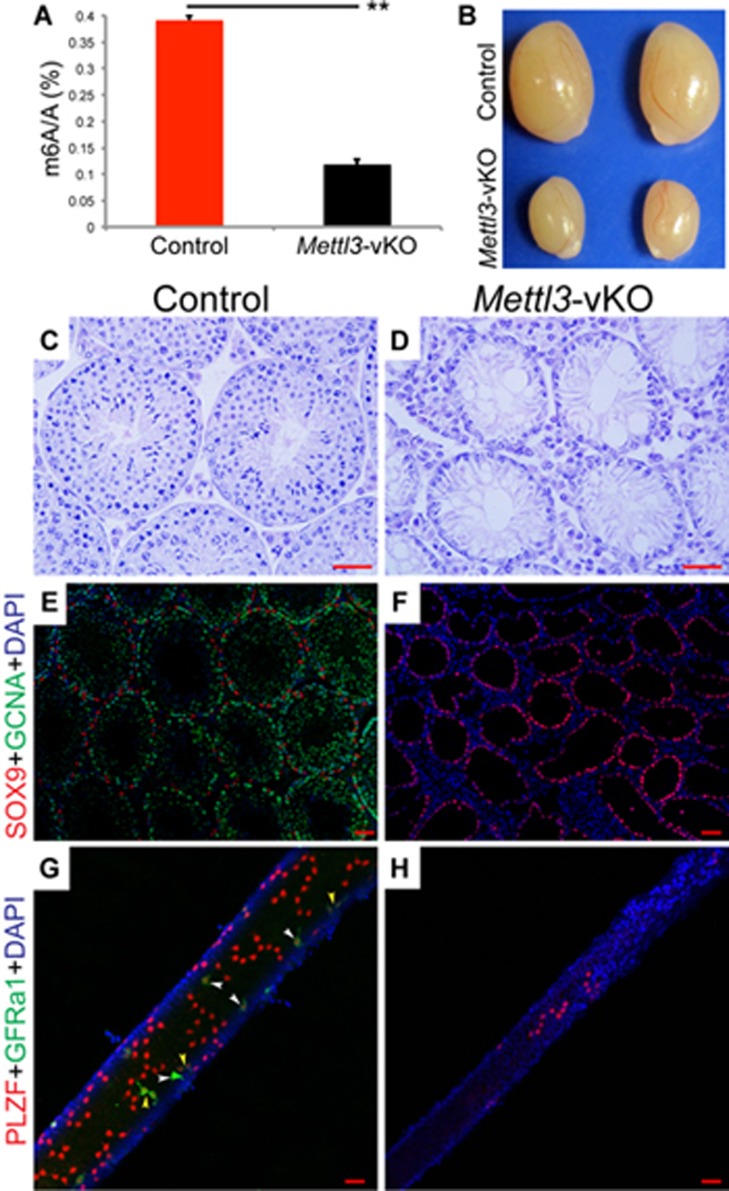
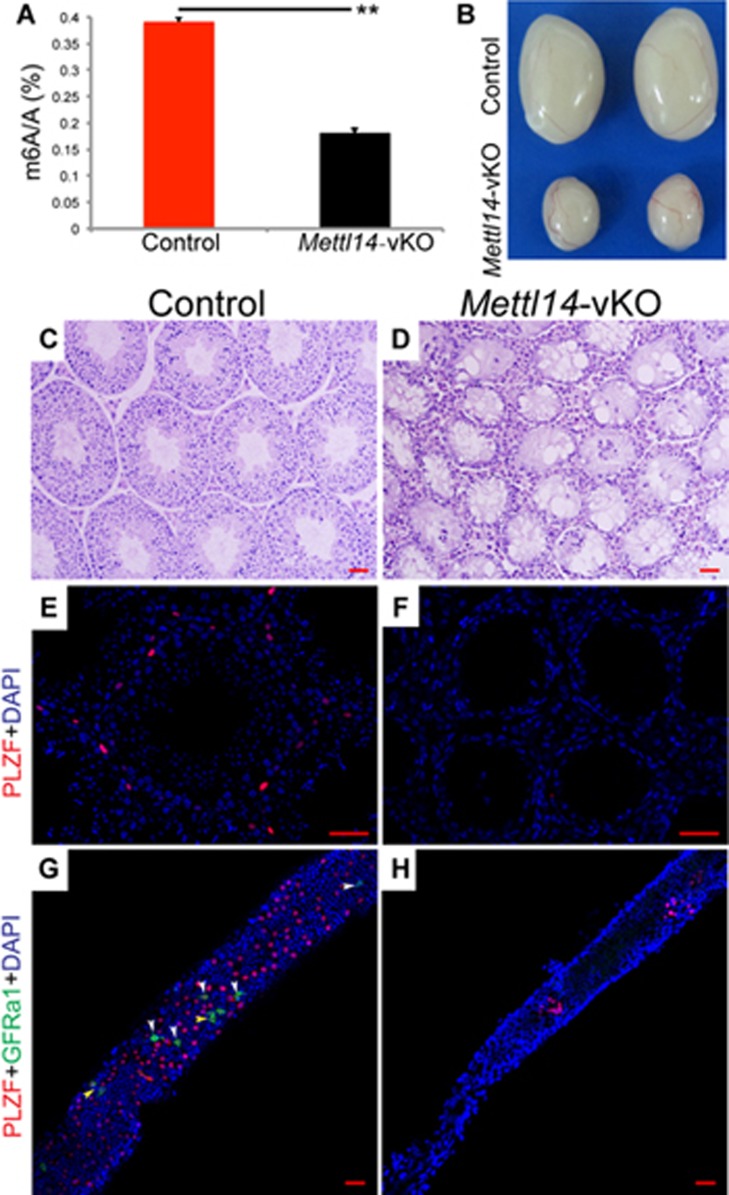
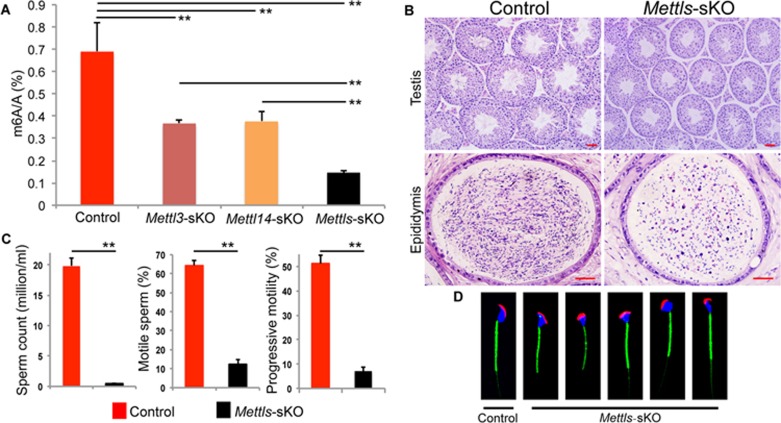
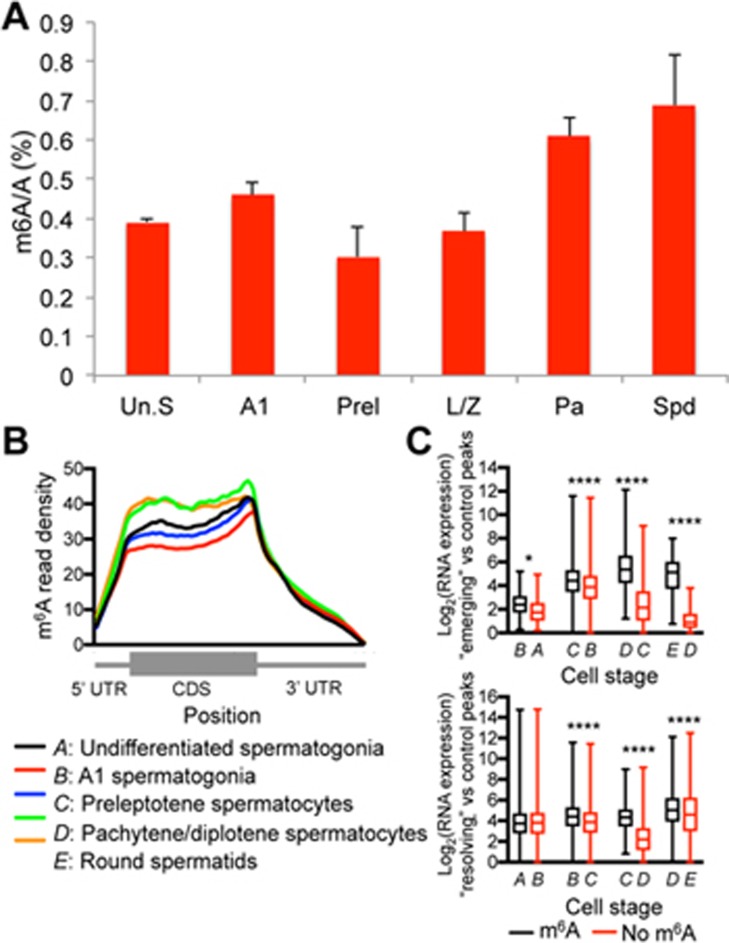
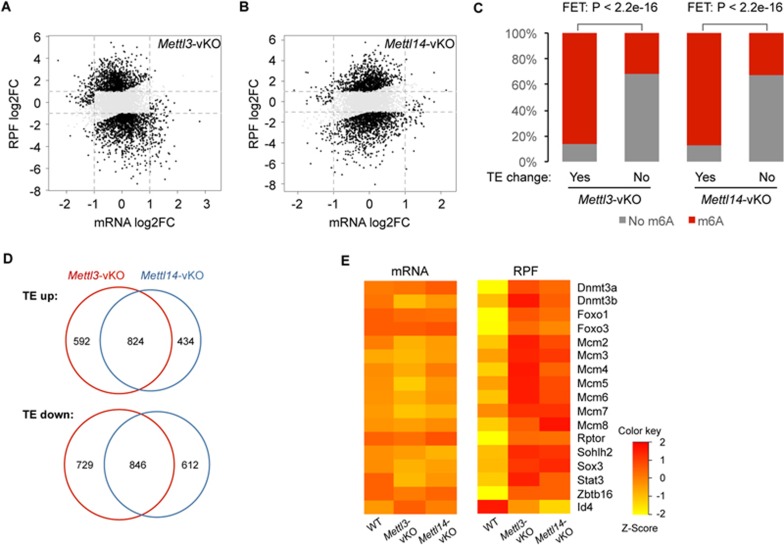
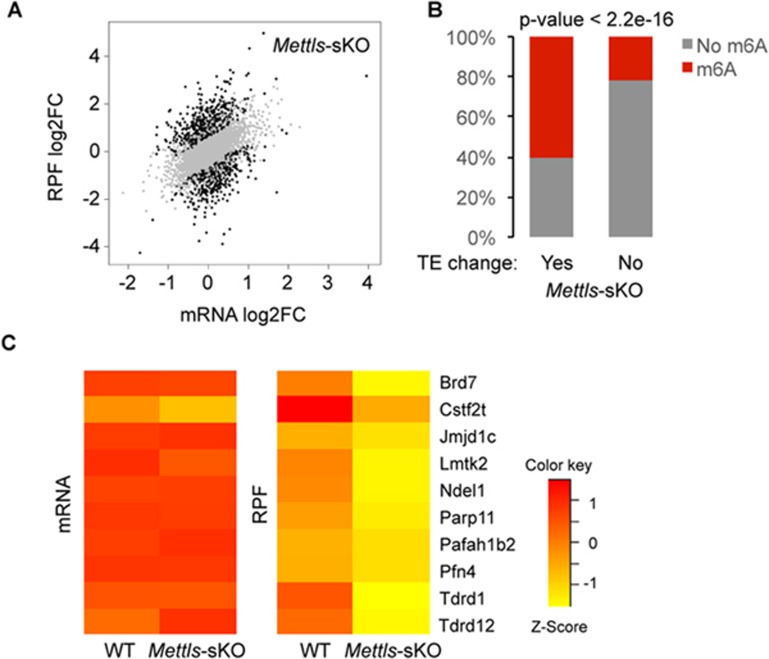
Spermatogenesis is a differentiation process during which diploid spermatogonial stem cells (SSCs) produce haploid spermatozoa. This highly specialized process is precisely controlled at the transcriptional, posttranscriptional, and translational levels. Here we report that N6-methyladenosine (m6A), an epitranscriptomic mark regulating gene expression, plays essential roles during spermatogenesis. We present comprehensive m6A mRNA methylomes of mouse spermatogenic cells from five developmental stages: undifferentiated spermatogonia, type A1 spermatogonia, preleptotene spermatocytes, pachytene/diplotene spermatocytes, and round spermatids. Germ cell-specific inactivation of the m6A RNA methyltransferase Mettl3 or Mettl14 with Vasa-Cre causes loss of m6A and depletion of SSCs. m6A depletion dysregulates translation of transcripts that are required for SSC proliferation/differentiation. Combined deletion of Mettl3 and Mettl14 in advanced germ cells with Stra8-GFPCre disrupts spermiogenesis, whereas mice with single deletion of either Mettl3 or Mettl14 in advanced germ cells show normal spermatogenesis. The spermatids from double-mutant mice exhibit impaired translation of haploid-specific genes that are essential for spermiogenesis. This study highlights crucial roles of mRNA m6A modification in germline development, potentially ensuring coordinated translation at different stages of spermatogenesis.
Figures
Similar articles
-
Periodic production of retinoic acid by meiotic and somatic cells coordinates four transitions in mouse spermatogenesis.Proc Natl Acad Sci U S A. 2017 Nov 21;114(47):E10132-E10141. doi: 10.1073/pnas.1710837114. Epub 2017 Nov 6. Proc Natl Acad Sci U S A. 2017. PMID: 29109271 Free PMC article.
-
[The Roles of N6-Methyladenosine Modification and Its Regulators in Male Reproduction].Sichuan Da Xue Xue Bao Yi Xue Ban. 2024 May 20;55(3):527-534. doi: 10.12182/20240560103. Sichuan Da Xue Xue Bao Yi Xue Ban. 2024. PMID: 38948273 Free PMC article. Review. Chinese.
-
Gene expression profiles in different stages of mouse spermatogenic cells during spermatogenesis.Biol Reprod. 2003 Jul;69(1):37-47. doi: 10.1095/biolreprod.102.012609. Epub 2003 Feb 19. Biol Reprod. 2003. PMID: 12606389
-
Transcriptome-wide Dynamics of m6A mRNA Methylation During Porcine Spermatogenesis.Genomics Proteomics Bioinformatics. 2023 Aug;21(4):729-741. doi: 10.1016/j.gpb.2021.08.006. Epub 2021 Sep 17. Genomics Proteomics Bioinformatics. 2023. PMID: 34543723 Free PMC article.
-
Mechanisms Regulating Spermatogonial Differentiation.Results Probl Cell Differ. 2016;58:253-87. doi: 10.1007/978-3-319-31973-5_10. Results Probl Cell Differ. 2016. PMID: 27300182 Review.
Cited by
-
Epigenetic regulations in mammalian spermatogenesis: RNA-m6A modification and beyond.Cell Mol Life Sci. 2021 Jun;78(11):4893-4905. doi: 10.1007/s00018-021-03823-9. Epub 2021 Apr 9. Cell Mol Life Sci. 2021. PMID: 33835194 Free PMC article. Review.
-
F1 Male Sterility in Cattle-Yak Examined through Changes in Testis Tissue and Transcriptome Profiles.Animals (Basel). 2022 Oct 9;12(19):2711. doi: 10.3390/ani12192711. Animals (Basel). 2022. PMID: 36230452 Free PMC article.
-
Mettl3‑mediated m6A RNA methylation regulates osteolysis induced by titanium particles.Mol Med Rep. 2024 Mar;29(3):36. doi: 10.3892/mmr.2024.13160. Epub 2024 Jan 12. Mol Med Rep. 2024. PMID: 38214327 Free PMC article.
-
m6A modification in RNA: biogenesis, functions and roles in gliomas.J Exp Clin Cancer Res. 2020 Sep 17;39(1):192. doi: 10.1186/s13046-020-01706-8. J Exp Clin Cancer Res. 2020. PMID: 32943100 Free PMC article. Review.
-
PCAF-mediated acetylation of METTL3 impairs mRNA translation efficiency in response to oxidative stress.Sci China Life Sci. 2024 Oct;67(10):2157-2168. doi: 10.1007/s11427-023-2535-x. Epub 2024 Aug 3. Sci China Life Sci. 2024. PMID: 39096338
References
-
- Clermont Y. Kinetics of spermatogenesis in mammals: seminiferous epithelium cycle and spermatogonial renewal. Physiol Rev 1972; 52: 198–236. - PubMed
-
- Kleene KC. Connecting cis-elements and trans-factors with mechanisms of developmental regulation of mRNA translation in meiotic and haploid mammalian spermatogenic cells. Reproduction 2013; 146:R1–R19. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
