

Hepatic inflammation caused by dysregulated bile acid synthesis is reversible by butyrate supplementation
- PMID: 28892150
- PMCID: PMC5953422
- DOI: 10.1002/path.4983
Hepatic inflammation caused by dysregulated bile acid synthesis is reversible by butyrate supplementation
Abstract
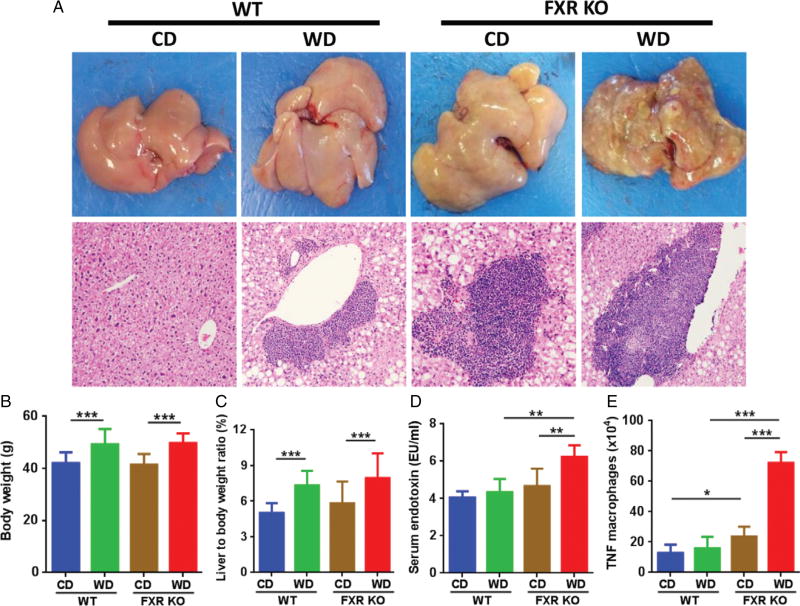
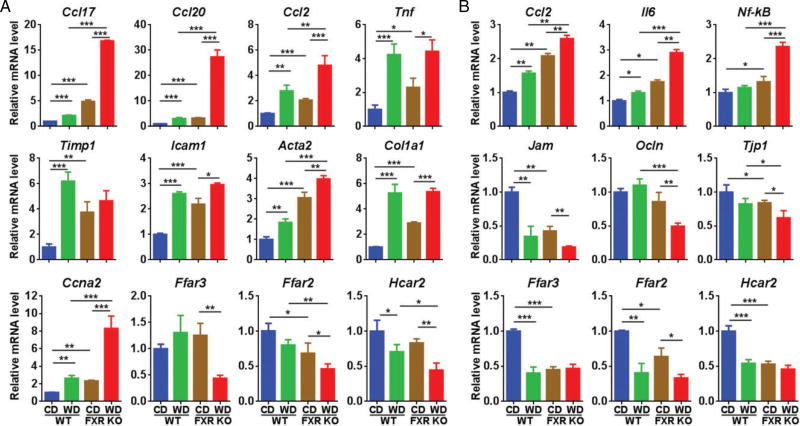
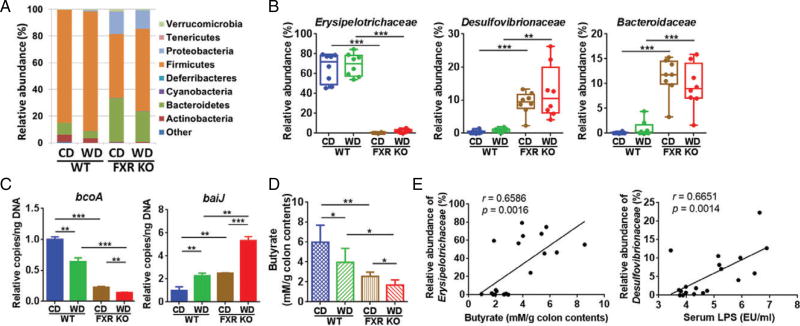
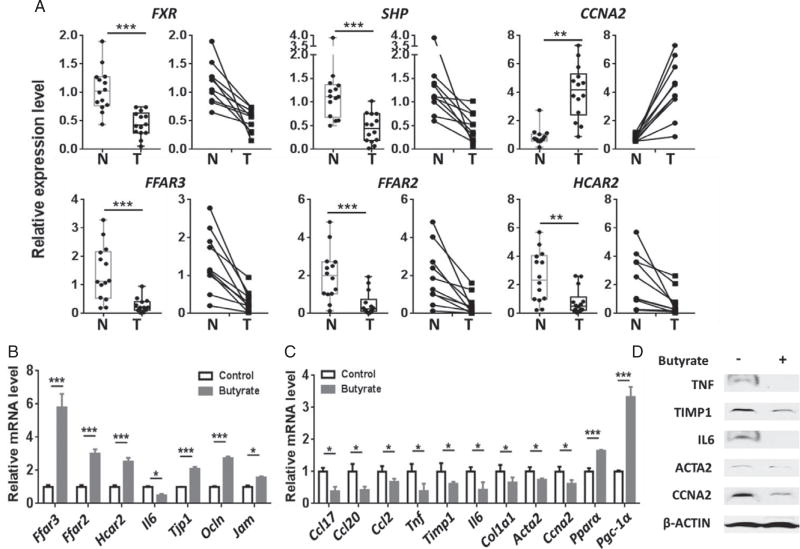
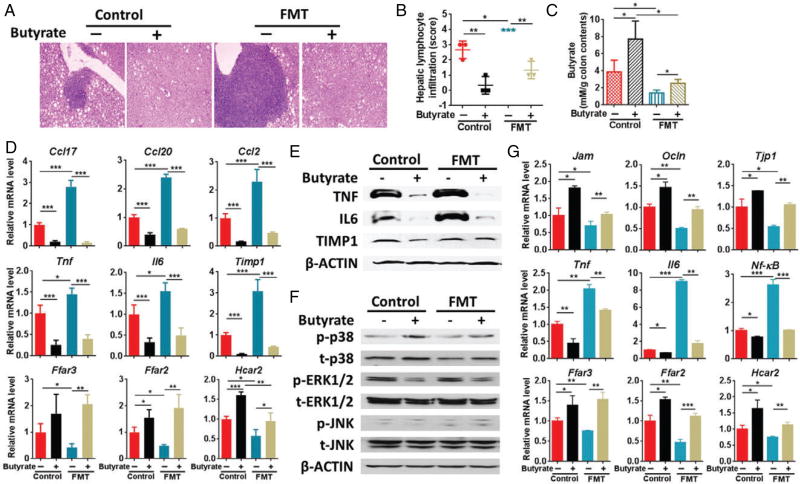
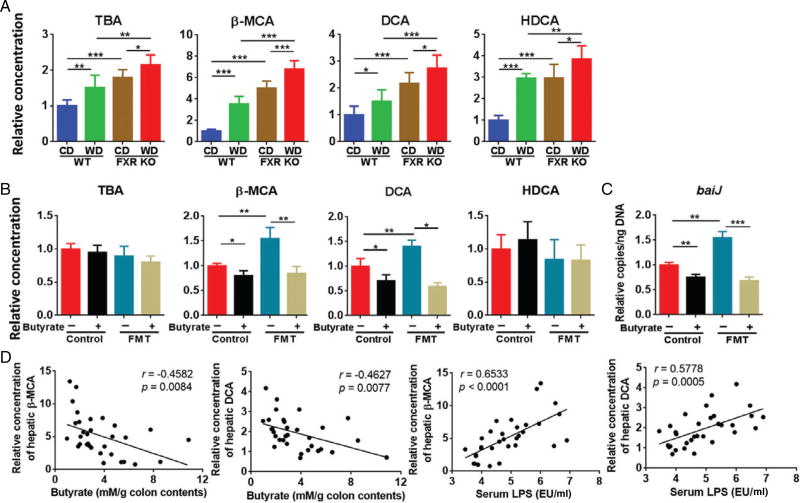
Dysregulated bile acid (BA) synthesis or reduced farnesoid X receptor (FXR) levels are found in patients having metabolic diseases, autoimmune hepatitis, and liver cirrhosis or cancer. The objective of this study was to establish the relationship between butyrate and dysregulated BA synthesis-induced hepatitis as well as the effect of butyrate in reversing the liver pathology. Wild-type (WT) and FXR knockout (KO) male mice were placed on a control (CD) or western diet (WD) for 15 months. In the presence or absence of butyrate supplementation, feces obtained from 15-month-old WD-fed FXR KO mice, which had severe hepatitis and liver tumors, were transplanted to 7-month-old WD-fed FXR KO for 3 months. Hepatic phenotypes, microbiota profile, and BA composition were analyzed. Butyrate-generating bacteria and colonic butyrate concentration were reduced due to FXR inactivation and further reduced by WD intake. In addition, WD-fed FXR KO male mice had the highest concentration of hepatic β-muricholic acid (β-MCA) and bacteria-generated deoxycholic acid (DCA) accompanied by serious hepatitis. Moreover, dysregulated BA and reduced SCFA signaling co-existed in both human liver cancers and WD-fed FXR KO mice. Microbiota transplantation using butyrate-deficient feces derived from 15-month-old WD-fed FXR KO mice increased hepatic lymphocyte numbers as well as hepatic β-MCA and DCA concentrations. Furthermore, butyrate supplementation reduced hepatic β-MCA as well as DCA and eliminated hepatic lymphocyte infiltration. In conclusion, reduced butyrate contributes to the development of hepatitis in the FXR KO mouse model. In addition, butyrate reverses dysregulated BA synthesis and its associated hepatitis. Copyright © 2017 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Keywords: FXR; gut microbiota; hepatitis; liver cancer; probiotics; short-chain fatty acids.
Copyright © 2017 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Conflict of interest statement
No conflicts of interest were declared.
Figures
Similar articles
-
Western Diet-Induced Dysbiosis in Farnesoid X Receptor Knockout Mice Causes Persistent Hepatic Inflammation after Antibiotic Treatment.Am J Pathol. 2017 Aug;187(8):1800-1813. doi: 10.1016/j.ajpath.2017.04.019. Epub 2017 Jul 12. Am J Pathol. 2017. PMID: 28711154 Free PMC article.
-
Gender Differences in Bile Acids and Microbiota in Relationship with Gender Dissimilarity in Steatosis Induced by Diet and FXR Inactivation.Sci Rep. 2017 May 11;7(1):1748. doi: 10.1038/s41598-017-01576-9. Sci Rep. 2017. PMID: 28496104 Free PMC article.
-
Dysregulated FXR-FGF19 signaling and choline metabolism are associated with gut dysbiosis and hyperplasia in a novel pig model of pediatric NASH.Am J Physiol Gastrointest Liver Physiol. 2020 Mar 1;318(3):G582-G609. doi: 10.1152/ajpgi.00344.2019. Epub 2020 Jan 31. Am J Physiol Gastrointest Liver Physiol. 2020. PMID: 32003601 Free PMC article.
-
The role of farnesoid X receptor in metabolic diseases, and gastrointestinal and liver cancer.Nat Rev Gastroenterol Hepatol. 2021 May;18(5):335-347. doi: 10.1038/s41575-020-00404-2. Epub 2021 Feb 10. Nat Rev Gastroenterol Hepatol. 2021. PMID: 33568795 Review.
-
An Intestinal Microbiota-Farnesoid X Receptor Axis Modulates Metabolic Disease.Gastroenterology. 2016 Nov;151(5):845-859. doi: 10.1053/j.gastro.2016.08.057. Epub 2016 Sep 14. Gastroenterology. 2016. PMID: 27639801 Free PMC article. Review.
Cited by
-
Gut Bacteria in Alcohol-Associated Liver Disease.Clin Liver Dis. 2024 Nov;28(4):663-679. doi: 10.1016/j.cld.2024.06.008. Epub 2024 Jul 24. Clin Liver Dis. 2024. PMID: 39362714 Review.
-
FGF21-dependent alleviation of cholestasis-induced liver fibrosis by sodium butyrate.Front Pharmacol. 2024 Jul 8;15:1422770. doi: 10.3389/fphar.2024.1422770. eCollection 2024. Front Pharmacol. 2024. PMID: 39040469 Free PMC article.
-
Exploring Advanced Therapies for Primary Biliary Cholangitis: Insights from the Gut Microbiota-Bile Acid-Immunity Network.Int J Mol Sci. 2024 Apr 13;25(8):4321. doi: 10.3390/ijms25084321. Int J Mol Sci. 2024. PMID: 38673905 Free PMC article. Review.
-
Effects of Bacillus Calmette-Guérin on immunometabolism, microbiome and liver diseases⋆.Liver Res. 2023 Jun;7(2):116-123. doi: 10.1016/j.livres.2023.05.001. Epub 2023 May 19. Liver Res. 2023. PMID: 38223885 Free PMC article.
-
The gut-liver axis in hepatobiliary diseases.Inflamm Regen. 2024 Jan 8;44(1):2. doi: 10.1186/s41232-023-00315-0. Inflamm Regen. 2024. PMID: 38191517 Free PMC article. Review.
References
-
- Lefebvre P, Cariou B, Lien F, et al. Role of bile acids and bile acid receptors in metabolic regulation. Physiol Rev. 2009;89:147–191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
