

Lyn and Fyn function as molecular switches that control immunoreceptors to direct homeostasis or inflammation
- PMID: 28811476
- PMCID: PMC5557797
- DOI: 10.1038/s41467-017-00294-0
Lyn and Fyn function as molecular switches that control immunoreceptors to direct homeostasis or inflammation
Abstract
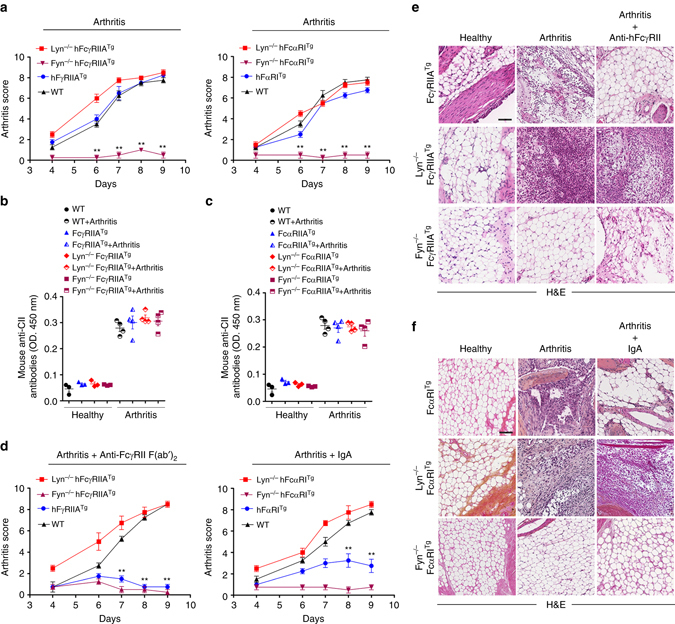
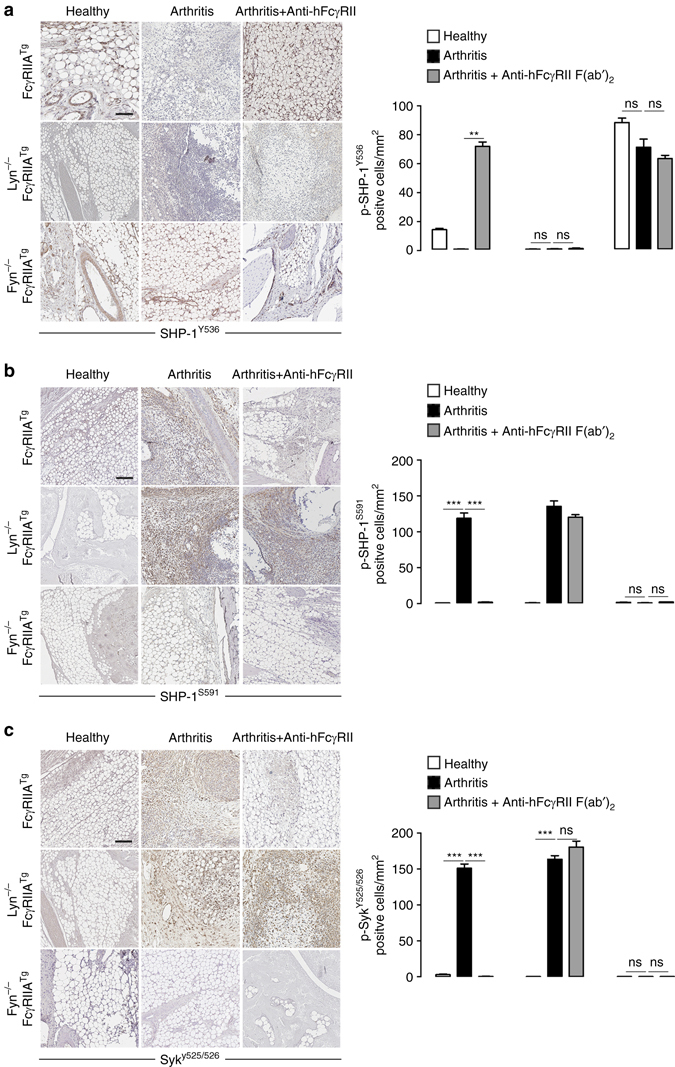
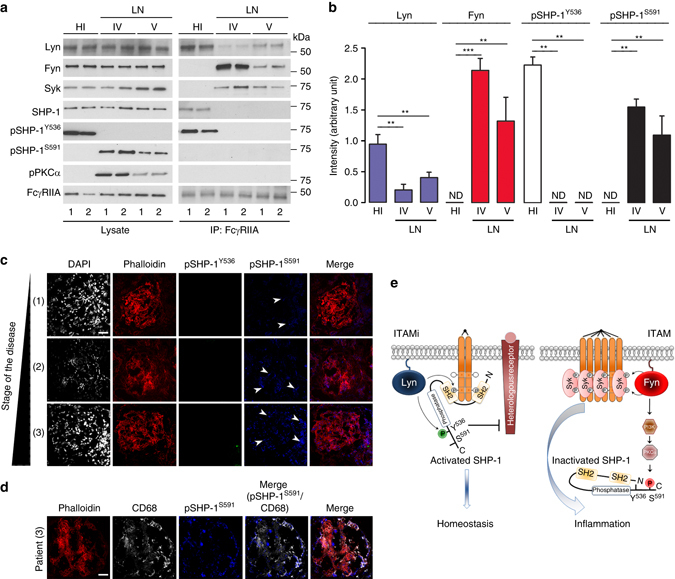
Immunoreceptors can transduce either inhibitory or activatory signals depending on ligand avidity and phosphorylation status, which is modulated by the protein kinases Lyn and Fyn. Here we show that Lyn and Fyn control immune receptor signaling status. SHP-1 tyrosine 536 phosphorylation by Lyn activates the phosphatase promoting inhibitory signaling through the immunoreceptor. By contrast, Fyn-dependent phosphorylation of SHP-1 serine 591 inactivates the phosphatase, enabling activatory immunoreceptor signaling. These SHP-1 signatures are relevant in vivo, as Lyn deficiency exacerbates nephritis and arthritis in mice, whereas Fyn deficiency is protective. Similarly, Fyn-activating signature is detected in patients with lupus nephritis, underlining the importance of this Lyn-Fyn balance. These data show how receptors discriminate negative from positive signals that respectively result in homeostatic or inflammatory conditions.Src-family kinases Fyn and Lyn are signaling components downstream of ITAM-bearing antigen receptors. Here the authors show that by phosphorylating SHP-1 at different residues, Lyn and Fyn can have opposing regulatory effects on ITAM receptors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
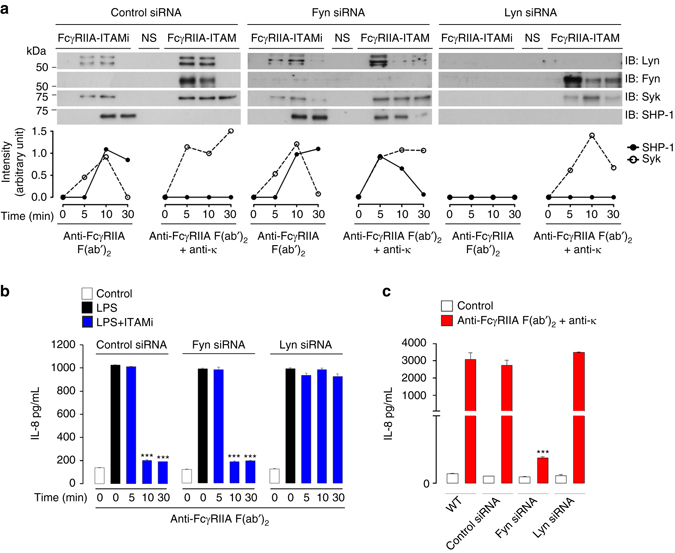
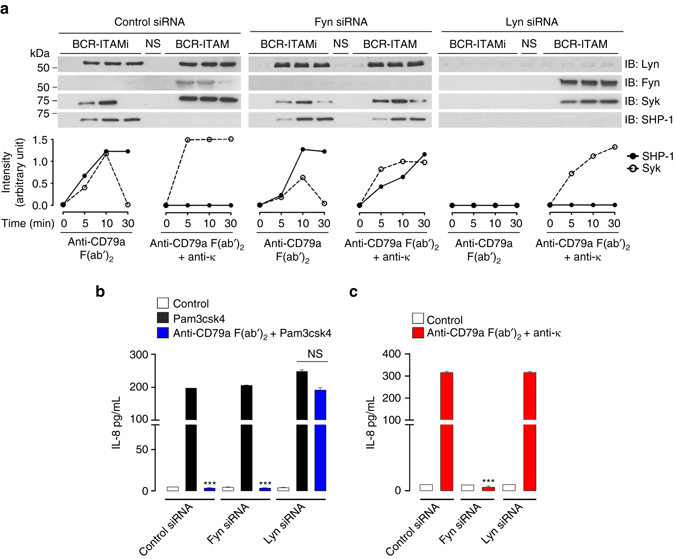
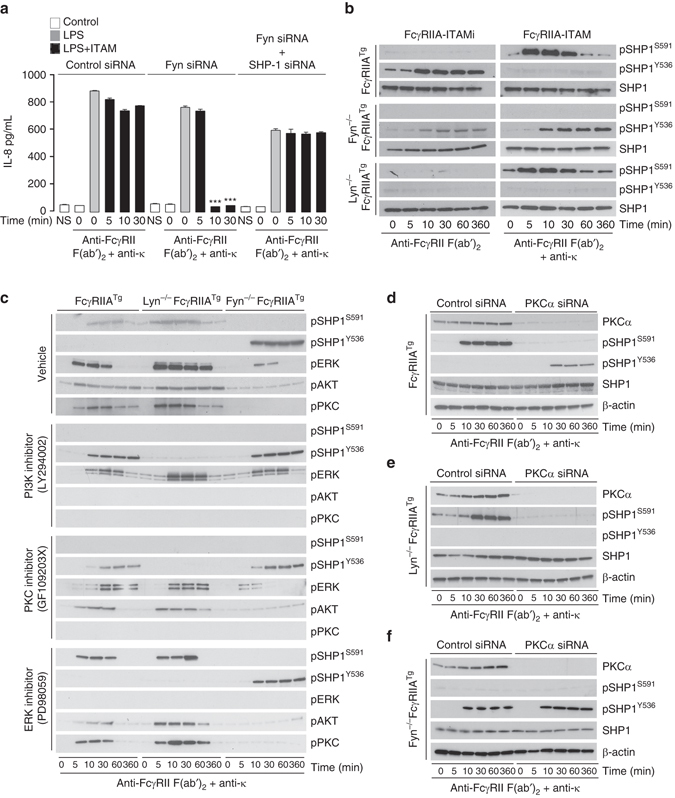
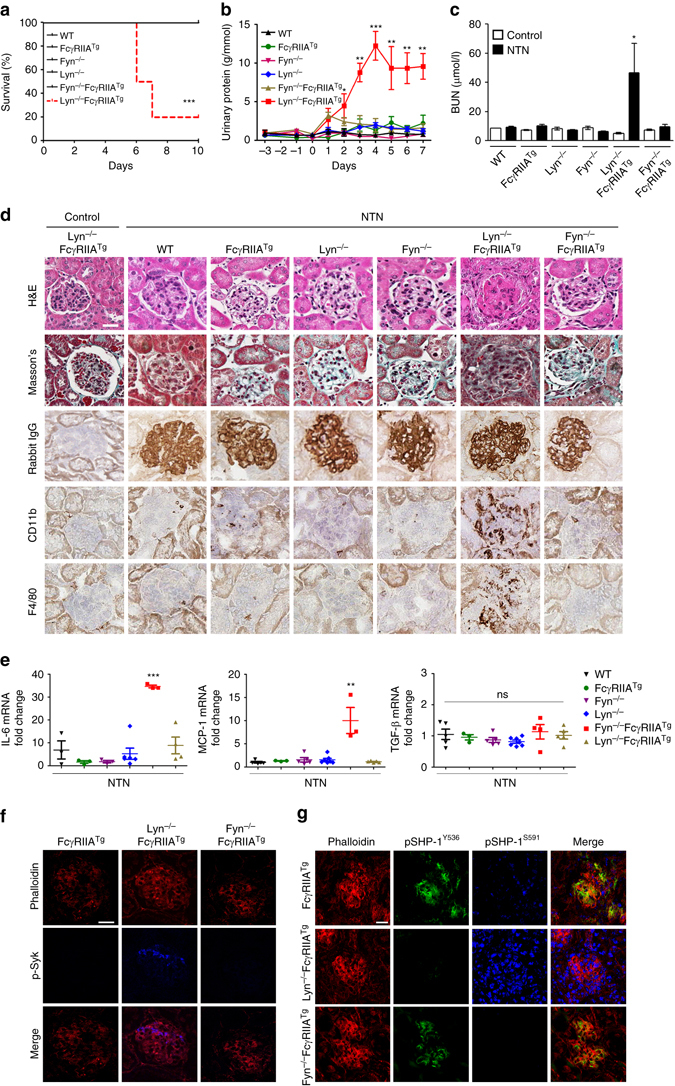
Comment in
-
Immunology: Balancing immunoreceptor signalling.Nat Rev Rheumatol. 2017 Oct;13(10):570. doi: 10.1038/nrrheum.2017.149. Epub 2017 Aug 31. Nat Rev Rheumatol. 2017. PMID: 28855691 No abstract available.
Similar articles
-
The role of Src family kinases in mast cell effector function.Novartis Found Symp. 2005;271:39-47; discussion 47-53, 95-9. Novartis Found Symp. 2005. PMID: 16605127 Review.
-
PTPalpha activates Lyn and Fyn and suppresses Hck to negatively regulate FcepsilonRI-dependent mast cell activation and allergic responses.J Immunol. 2010 Nov 15;185(10):5993-6002. doi: 10.4049/jimmunol.1001261. Epub 2010 Oct 13. J Immunol. 2010. PMID: 20944008
-
Lyn but not Fyn kinase controls IgG-mediated systemic anaphylaxis.J Immunol. 2012 May 1;188(9):4360-8. doi: 10.4049/jimmunol.1003223. Epub 2012 Mar 26. J Immunol. 2012. PMID: 22450804 Free PMC article.
-
Regulation of the tyrosine phosphorylation of Phospholipid Scramblase 1 in mast cells that are stimulated through the high-affinity IgE receptor.PLoS One. 2014 Oct 7;9(10):e109800. doi: 10.1371/journal.pone.0109800. eCollection 2014. PLoS One. 2014. PMID: 25289695 Free PMC article.
-
Signaling mutations and autoimmunity.Curr Dir Autoimmun. 2003;6:61-88. doi: 10.1159/000066856. Curr Dir Autoimmun. 2003. PMID: 12408047 Review.
Cited by
-
Integrative analysis of macrophage ribo-Seq and RNA-Seq data define glucocorticoid receptor regulated inflammatory response genes into distinct regulatory classes.Comput Struct Biotechnol J. 2022 Oct 3;20:5622-5638. doi: 10.1016/j.csbj.2022.09.042. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36284713 Free PMC article.
-
Proteomic analysis of plasma exosomes from Cystic Echinococcosis patients provides in vivo support for distinct immune response profiles in active vs inactive infection and suggests potential biomarkers.PLoS Negl Trop Dis. 2020 Oct 5;14(10):e0008586. doi: 10.1371/journal.pntd.0008586. eCollection 2020 Oct. PLoS Negl Trop Dis. 2020. PMID: 33017416 Free PMC article.
-
MicroRNAs: Potential prognostic and theranostic biomarkers in chronic lymphocytic leukemia.EJHaem. 2024 Feb 11;5(1):191-205. doi: 10.1002/jha2.849. eCollection 2024 Feb. EJHaem. 2024. PMID: 38406506 Free PMC article. Review.
-
Editorial: The Role of Inhibitory Receptors in Inflammation and Cancer.Front Immunol. 2020 Dec 22;11:633686. doi: 10.3389/fimmu.2020.633686. eCollection 2020. Front Immunol. 2020. PMID: 33414792 Free PMC article. No abstract available.
-
Biosensors based on peptide exposure show single molecule conformations in live cells.Cell. 2021 Oct 28;184(22):5670-5685.e23. doi: 10.1016/j.cell.2021.09.026. Epub 2021 Oct 11. Cell. 2021. PMID: 34637702 Free PMC article.
References
-
- Hamerman JA, Lanier LL. Inhibition of immune responses by ITAM-bearing receptors. Sci. STKE. 2006;2006:re1. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
