

Interferon-Inducible CD169/Siglec1 Attenuates Anti-HIV-1 Effects of Alpha Interferon
- PMID: 28794041
- PMCID: PMC5640826
- DOI: 10.1128/JVI.00972-17
Interferon-Inducible CD169/Siglec1 Attenuates Anti-HIV-1 Effects of Alpha Interferon
Abstract
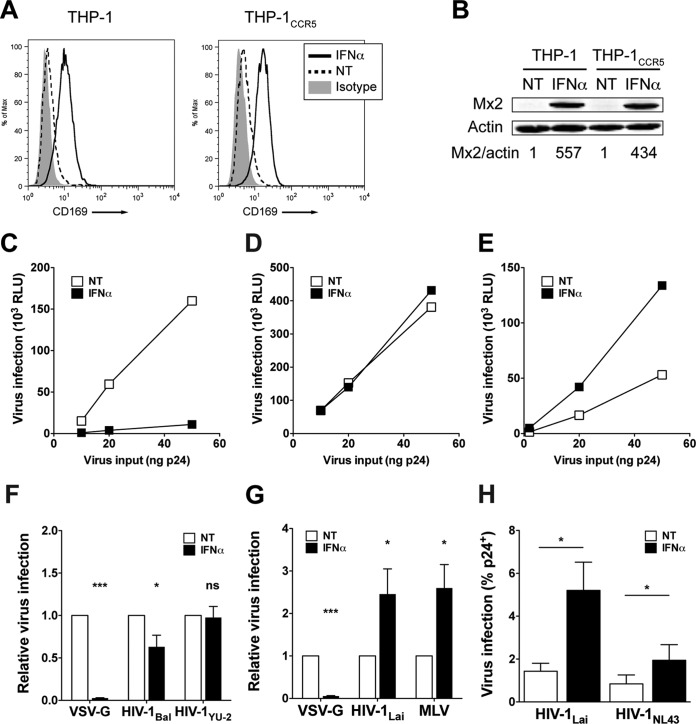
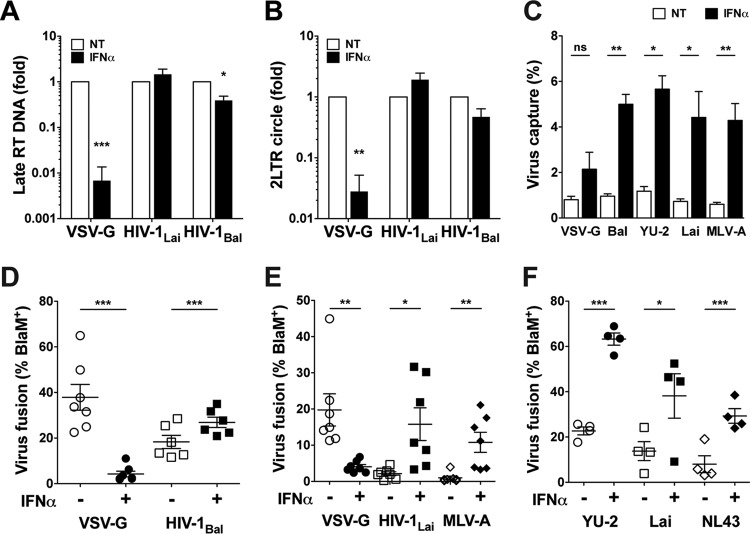
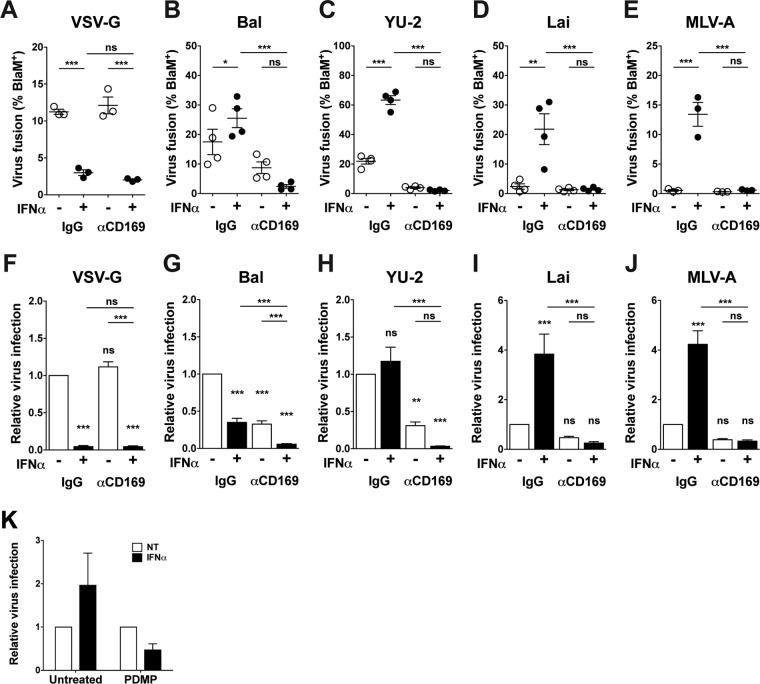
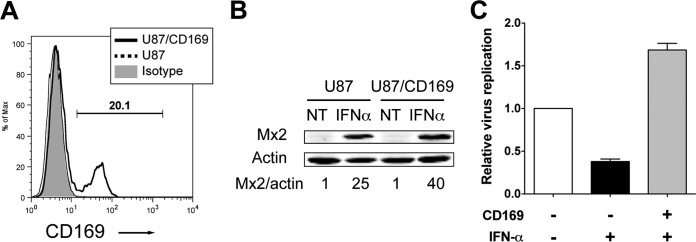
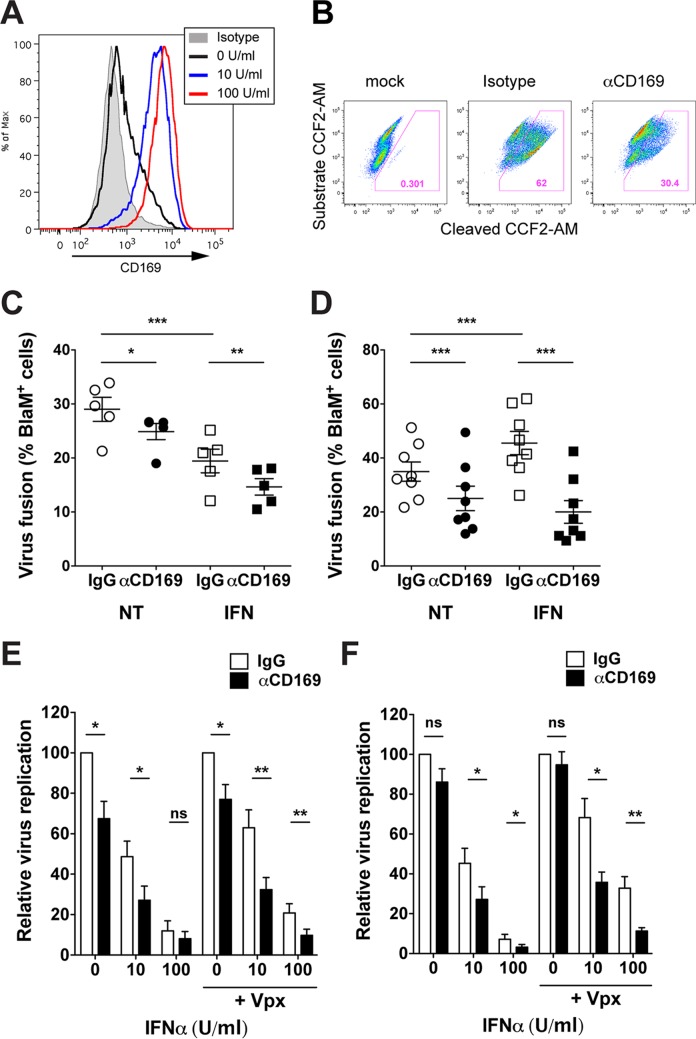
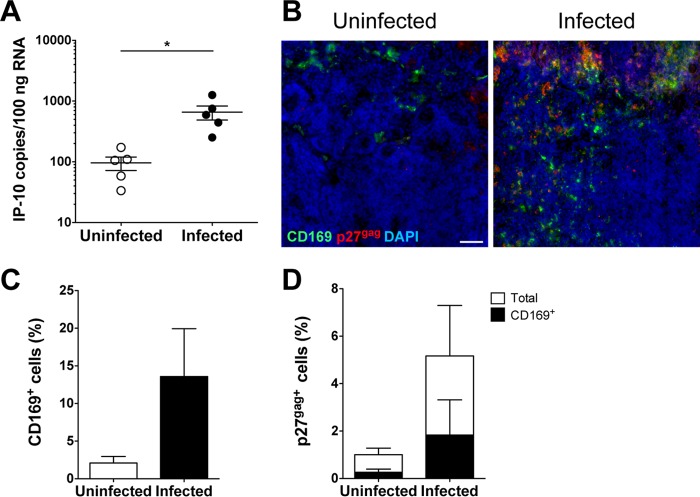
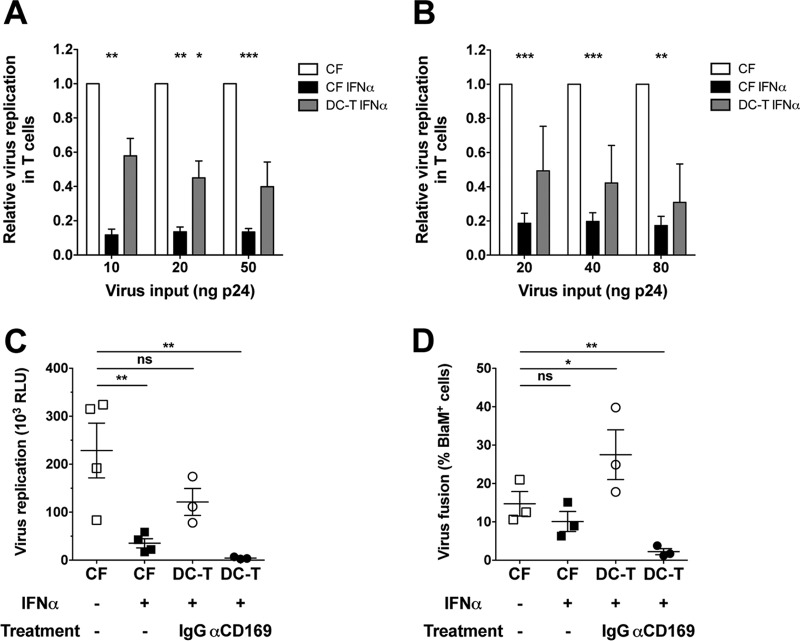
A hallmark of human immunodeficiency virus type 1 (HIV-1) infection in vivo is chronic immune activation concomitant with type I interferon (IFN) production. Although type I IFN induces an antiviral state in many cell types, HIV-1 can replicate in vivo via mechanisms that have remained unclear. We have recently identified a type I IFN-inducible protein, CD169, as the HIV-1 attachment factor on dendritic cells (DCs) that can mediate robust infection of CD4+ T cells in trans Since CD169 expression on macrophages is also induced by type I IFN, we hypothesized that type I IFN-inducible CD169 could facilitate productive HIV-1 infection in myeloid cells in cis and CD4+ T cells in trans and thus offset antiviral effects of type I IFN. In support of this hypothesis, infection of HIV-1 or murine leukemia virus Env (MLV-Env)-pseudotyped HIV-1 particles was enhanced in IFN-α-treated THP-1 monocytoid cells, and this enhancement was primarily dependent on CD169-mediated enhancement at the virus entry step, a phenomenon phenocopied in HIV-1 infections of IFN-α-treated primary monocyte-derived macrophages (MDMs). Furthermore, expression of CD169, a marker of type I IFN-induced immune activation in vivo, was enhanced in lymph nodes from pigtailed macaques infected with simian immunodeficiency virus (SIV) carrying HIV-1 reverse transcriptase (RT-SHIV), compared to uninfected macaques, and interestingly, there was extensive colocalization of p27gag and CD169, suggesting productive infection of CD169+ myeloid cells in vivo While cell-free HIV-1 infection of IFN-α-treated CD4+ T cells was robustly decreased, initiation of infection in trans via coculture with CD169+ IFN-α-treated DCs restored infection, suggesting that HIV-1 exploits CD169 in cis and in trans to attenuate a type I IFN-induced antiviral state.IMPORTANCE HIV-1 infection in humans causes immune activation characterized by elevated levels of proinflammatory cytokines, including type I interferons (IFN). Although type I IFN induces an antiviral state in many cell types in vitro, HIV-1 can replicate in vivo via mechanisms that have remained unclear. In this study, we tested the hypothesis that CD169, a type I IFN-inducible HIV-1 attachment factor, offsets antiviral effects of type I IFN. Infection of HIV-1 was rescued in IFN-α-treated myeloid cells via upregulation of CD169 and a subsequent increase in CD169-dependent virus entry. Furthermore, extensive colocalization of viral Gag and CD169 was observed in lymph nodes of infected pigtailed macaques, suggesting productive infection of CD169+ cells in vivo Treatment of dendritic cell (DC)-T cell cocultures with IFN-α upregulated CD169 expression on DCs and rescued HIV-1 infection of CD4+ T cells in trans, suggesting that HIV-1 exploits CD169 to attenuate type I IFN-induced restrictions.
Keywords: CD169; HIV; type I IFN; virus entry; virus evasion.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Interferon-inducible mechanism of dendritic cell-mediated HIV-1 dissemination is dependent on Siglec-1/CD169.PLoS Pathog. 2013;9(4):e1003291. doi: 10.1371/journal.ppat.1003291. Epub 2013 Apr 11. PLoS Pathog. 2013. PMID: 23593001 Free PMC article.
-
Blocking TLR7- and TLR9-mediated IFN-α production by plasmacytoid dendritic cells does not diminish immune activation in early SIV infection.PLoS Pathog. 2013;9(7):e1003530. doi: 10.1371/journal.ppat.1003530. Epub 2013 Jul 25. PLoS Pathog. 2013. PMID: 23935491 Free PMC article.
-
Attenuation of pathogenic immune responses during infection with human and simian immunodeficiency virus (HIV/SIV) by the tetracycline derivative minocycline.PLoS One. 2014 Apr 14;9(4):e94375. doi: 10.1371/journal.pone.0094375. eCollection 2014. PLoS One. 2014. PMID: 24732038 Free PMC article.
-
Interferon α subtypes in HIV infection.Cytokine Growth Factor Rev. 2018 Apr;40:13-18. doi: 10.1016/j.cytogfr.2018.02.002. Epub 2018 Feb 13. Cytokine Growth Factor Rev. 2018. PMID: 29475588 Review.
-
Role of mu-opioids as cofactors in human immunodeficiency virus type 1 disease progression and neuropathogenesis.J Neurovirol. 2011 Aug;17(4):291-302. doi: 10.1007/s13365-011-0037-2. Epub 2011 Jul 7. J Neurovirol. 2011. PMID: 21735315 Free PMC article. Review.
Cited by
-
Myeloid cell interferon responses correlate with clearance of SARS-CoV-2.Nat Commun. 2022 Feb 3;13(1):679. doi: 10.1038/s41467-022-28315-7. Nat Commun. 2022. PMID: 35115549 Free PMC article.
-
Investigation of type I interferon responses in ANCA-associated vasculitis.Sci Rep. 2021 Apr 15;11(1):8272. doi: 10.1038/s41598-021-87760-4. Sci Rep. 2021. PMID: 33859290 Free PMC article.
-
Immuno-modulating properties of Tulathromycin in porcine monocyte-derived macrophages infected with porcine reproductive and respiratory syndrome virus.PLoS One. 2019 Aug 23;14(8):e0221560. doi: 10.1371/journal.pone.0221560. eCollection 2019. PLoS One. 2019. PMID: 31442273 Free PMC article.
-
Interferon Alpha Favors Macrophage Infection by Visceral Leishmania Species Through Upregulation of Sialoadhesin Expression.Front Immunol. 2020 Jun 9;11:1113. doi: 10.3389/fimmu.2020.01113. eCollection 2020. Front Immunol. 2020. PMID: 32582193 Free PMC article.
-
A virus-packageable CRISPR screen identifies host factors mediating interferon inhibition of HIV.Elife. 2018 Dec 6;7:e39823. doi: 10.7554/eLife.39823. Elife. 2018. PMID: 30520725 Free PMC article.
References
-
- Stacey AR, Norris PJ, Qin L, Haygreen EA, Taylor E, Heitman J, Lebedeva M, DeCamp A, Li D, Grove D, Self SG, Borrow P. 2009. Induction of a striking systemic cytokine cascade prior to peak viremia in acute human immunodeficiency virus type 1 infection, in contrast to more modest and delayed responses in acute hepatitis B and C virus infections. J Virol 83:3719–3733. doi:10.1128/JVI.01844-08. - DOI - PMC - PubMed
-
- Beignon AS, McKenna K, Skoberne M, Manches O, DaSilva I, Kavanagh DG, Larsson M, Gorelick RJ, Lifson JD, Bhardwaj N. 2005. Endocytosis of HIV-1 activates plasmacytoid dendritic cells via Toll-like receptor-viral RNA interactions. J Clin Invest 115:3265–3275. doi:10.1172/JCI26032. - DOI - PMC - PubMed
-
- Fonteneau JF, Larsson M, Beignon AS, McKenna K, Dasilva I, Amara A, Liu YJ, Lifson JD, Littman DR, Bhardwaj N. 2004. Human immunodeficiency virus type 1 activates plasmacytoid dendritic cells and concomitantly induces the bystander maturation of myeloid dendritic cells. J Virol 78:5223–5232. doi:10.1128/JVI.78.10.5223-5232.2004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
