

Molecular Mechanisms of Synaptic Vesicle Priming by Munc13 and Munc18
- PMID: 28772123
- PMCID: PMC5747255
- DOI: 10.1016/j.neuron.2017.07.004
Molecular Mechanisms of Synaptic Vesicle Priming by Munc13 and Munc18
Abstract
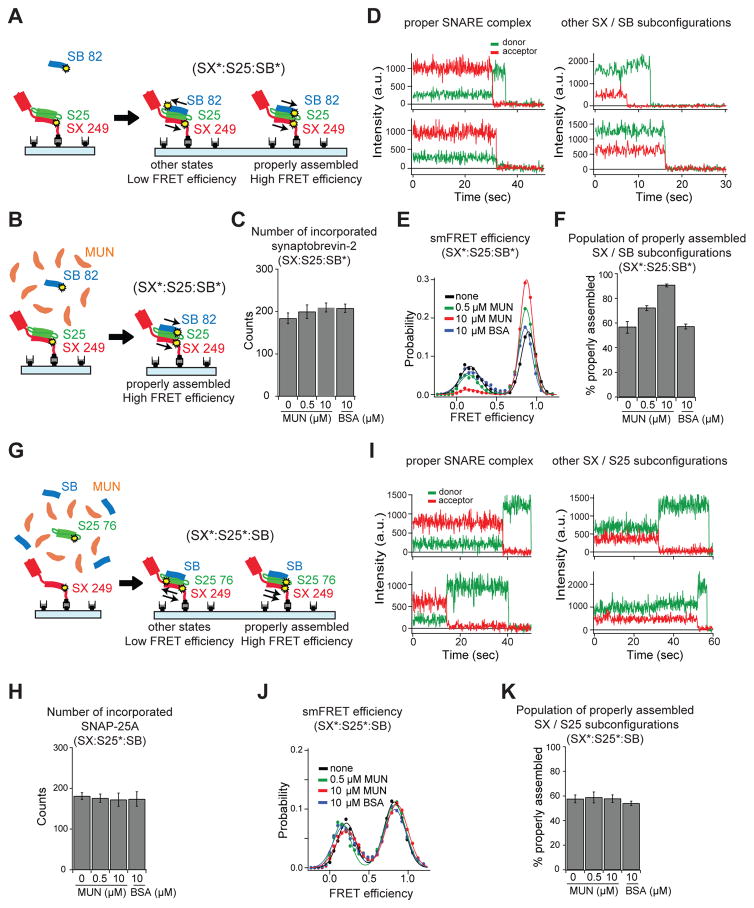
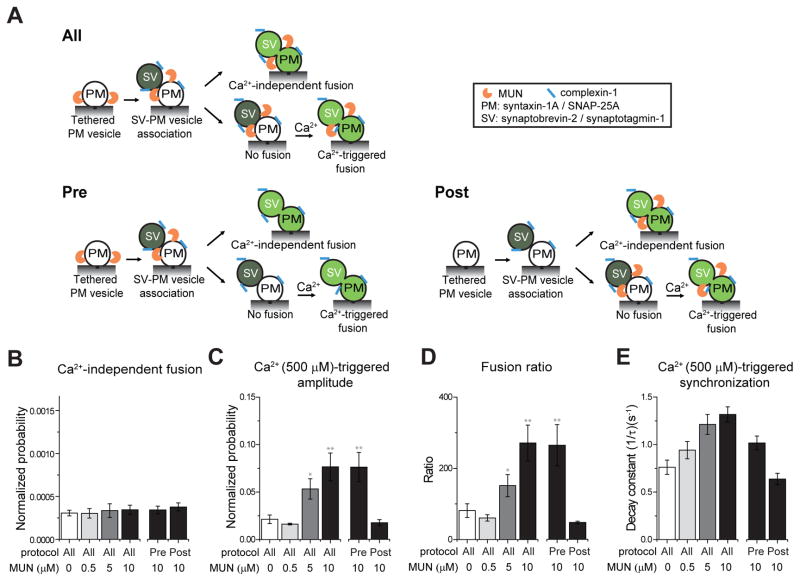
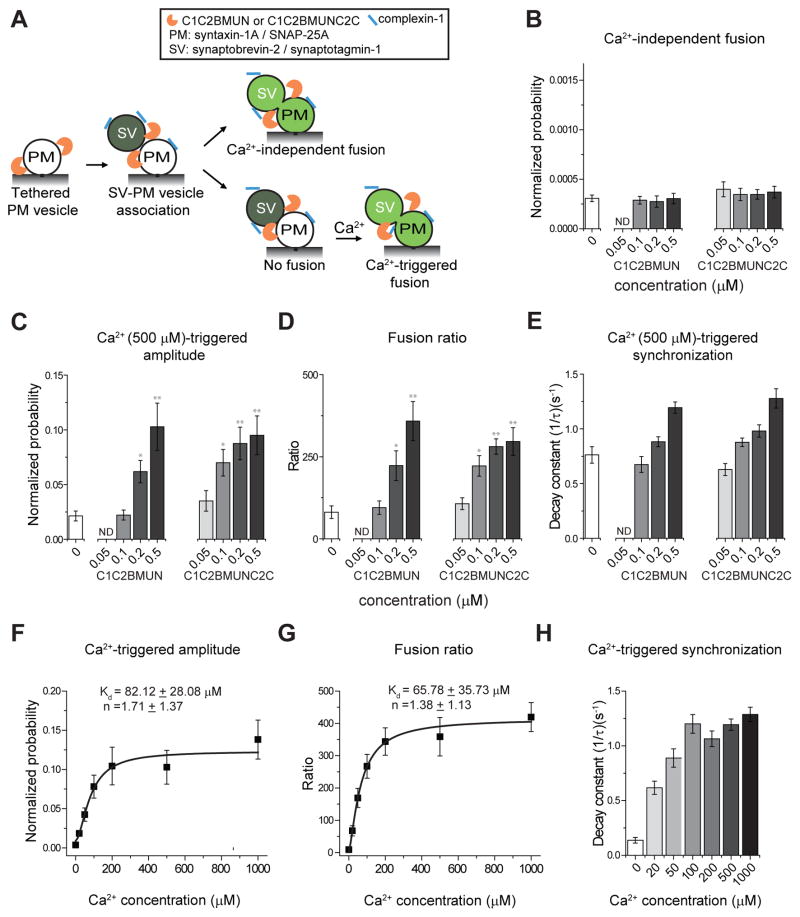
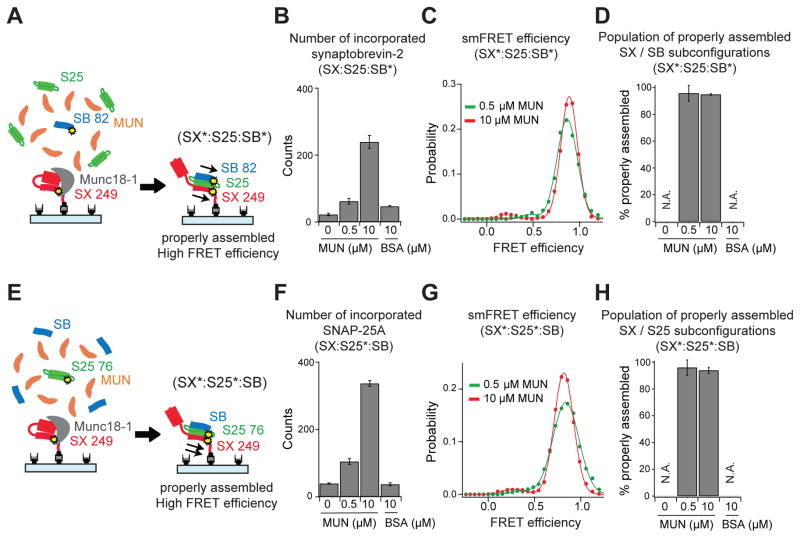
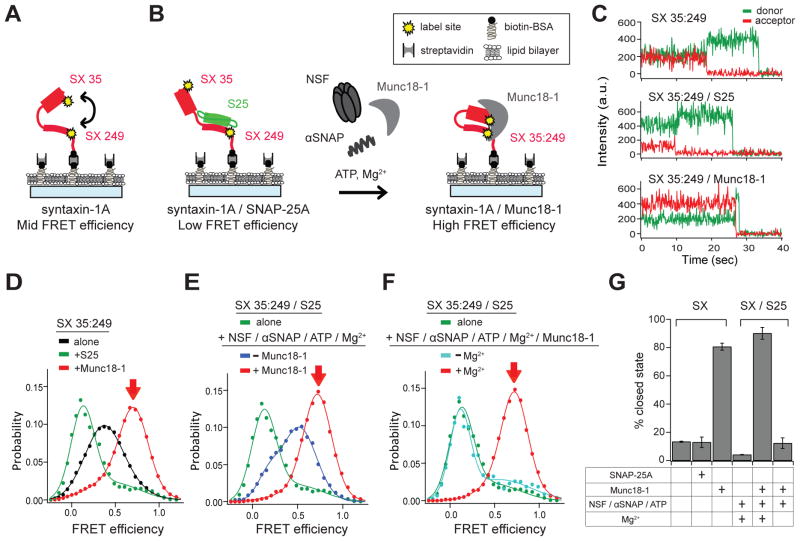
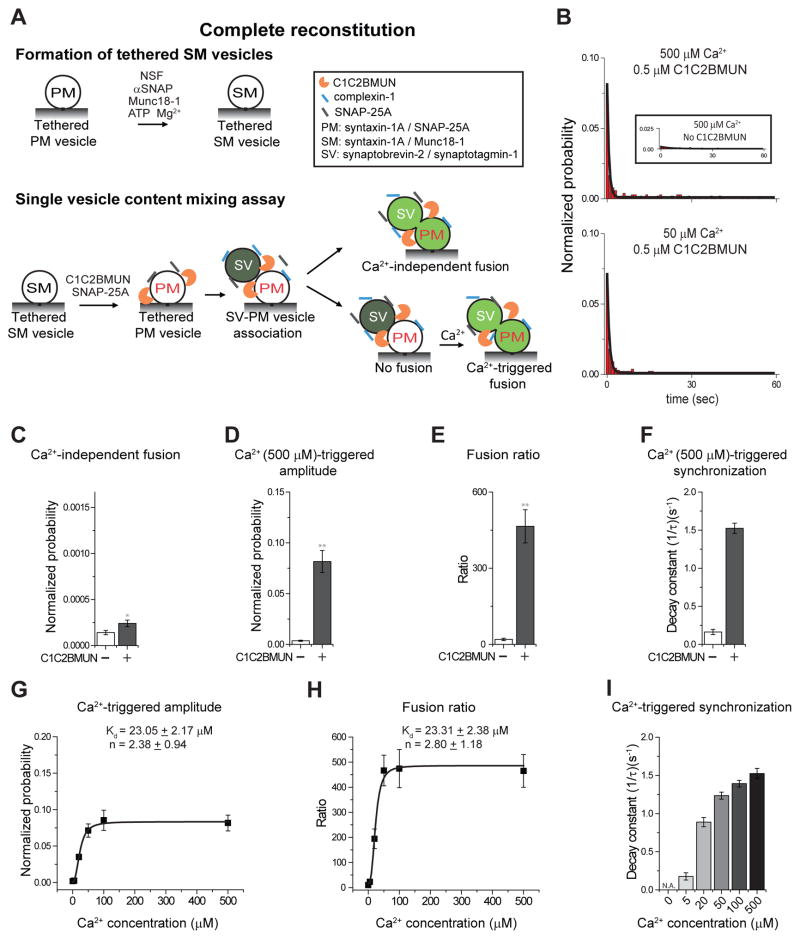
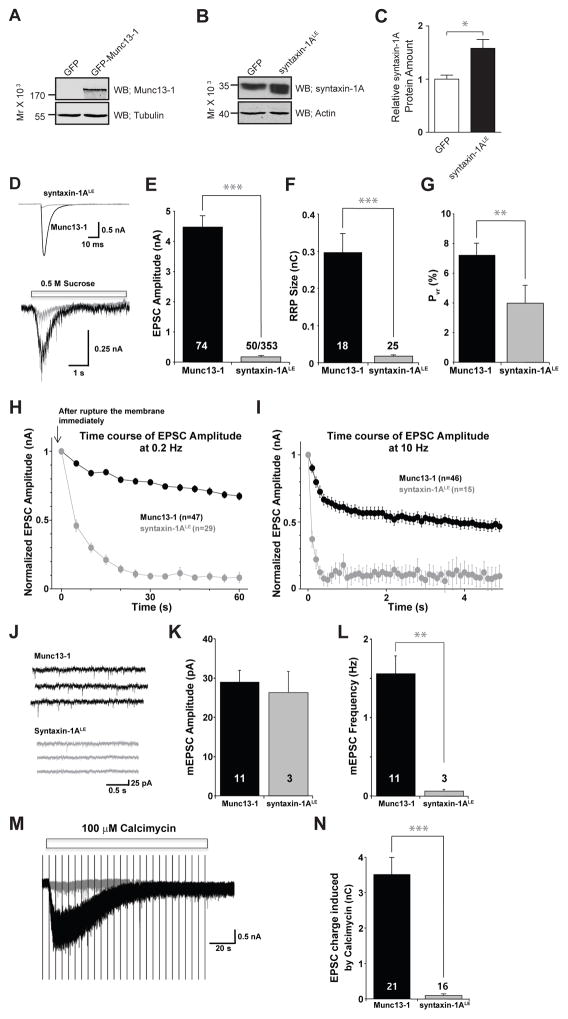
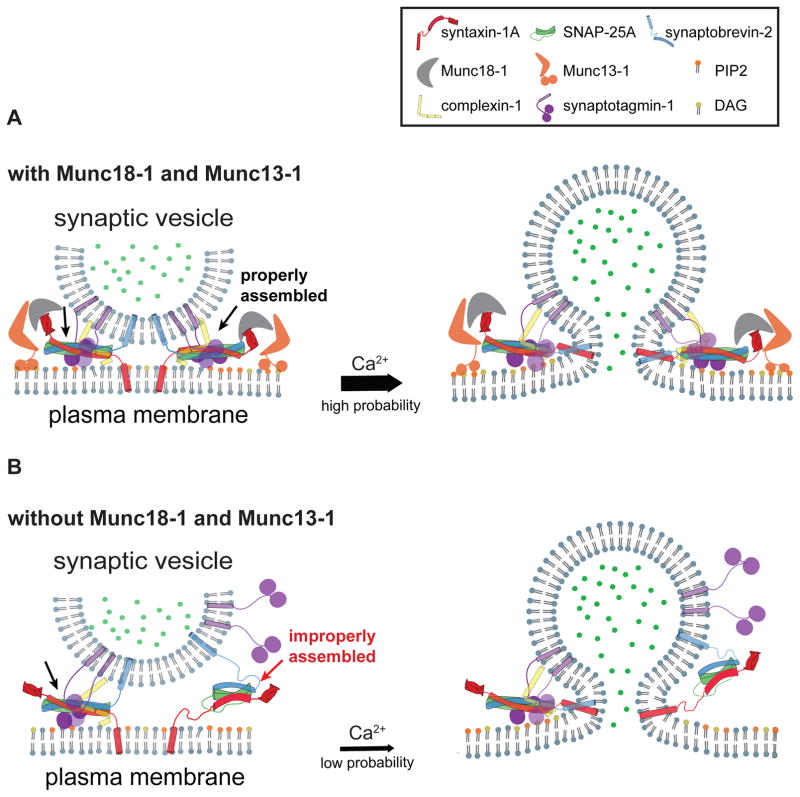
Munc13 catalyzes the transit of syntaxin from a closed complex with Munc18 into the ternary SNARE complex. Here we report a new function of Munc13, independent of Munc18: it promotes the proper syntaxin/synaptobrevin subconfiguration during assembly of the ternary SNARE complex. In cooperation with Munc18, Munc13 additionally ensures the proper syntaxin/SNAP-25 subconfiguration. In a reconstituted fusion assay with SNAREs, complexin, and synaptotagmin, inclusion of both Munc13 and Munc18 quadruples the Ca2+-triggered amplitude and achieves Ca2+ sensitivity at near-physiological concentrations. In Munc13-1/2 double-knockout neurons, expression of a constitutively open mutant of syntaxin could only minimally restore neurotransmitter release relative to Munc13-1 rescue. Together, the physiological functions of Munc13 may be related to regulation of proper SNARE complex assembly.
Keywords: Munc13; Munc18; complexin; neuronal SNAREs; neurotransmitter release; plasticity; priming; synaptotagmin.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Synaptotagmin-1-, Munc18-1-, and Munc13-1-dependent liposome fusion with a few neuronal SNAREs.Proc Natl Acad Sci U S A. 2021 Jan 26;118(4):e2019314118. doi: 10.1073/pnas.2019314118. Proc Natl Acad Sci U S A. 2021. PMID: 33468652 Free PMC article.
-
Munc13-1 MUN domain and Munc18-1 cooperatively chaperone SNARE assembly through a tetrameric complex.Proc Natl Acad Sci U S A. 2020 Jan 14;117(2):1036-1041. doi: 10.1073/pnas.1914361117. Epub 2019 Dec 30. Proc Natl Acad Sci U S A. 2020. PMID: 31888993 Free PMC article.
-
Reconstitution of the vital functions of Munc18 and Munc13 in neurotransmitter release.Science. 2013 Jan 25;339(6118):421-5. doi: 10.1126/science.1230473. Epub 2012 Dec 20. Science. 2013. PMID: 23258414 Free PMC article.
-
Neuronal SNARE complex assembly guided by Munc18-1 and Munc13-1.FEBS Open Bio. 2022 Nov;12(11):1939-1957. doi: 10.1002/2211-5463.13394. Epub 2022 Mar 22. FEBS Open Bio. 2022. PMID: 35278279 Free PMC article. Review.
-
The Synaptic Vesicle Release Machinery.Annu Rev Biophys. 2015;44:339-67. doi: 10.1146/annurev-biophys-060414-034057. Annu Rev Biophys. 2015. PMID: 26098518 Review.
Cited by
-
Control of Munc13-1 Activity by Autoinhibitory Interactions Involving the Variable N-terminal Region.bioRxiv [Preprint]. 2024 Jan 25:2024.01.24.577102. doi: 10.1101/2024.01.24.577102. bioRxiv. 2024. Update in: J Mol Biol. 2024 Apr 15;436(8):168502. doi: 10.1016/j.jmb.2024.168502. PMID: 38328168 Free PMC article. Updated. Preprint.
-
CaV1 and CaV2 calcium channels mediate the release of distinct pools of synaptic vesicles.Elife. 2023 Feb 23;12:e81407. doi: 10.7554/eLife.81407. Elife. 2023. PMID: 36820519 Free PMC article.
-
A sequential two-step priming scheme reproduces diversity in synaptic strength and short-term plasticity.Proc Natl Acad Sci U S A. 2022 Aug 23;119(34):e2207987119. doi: 10.1073/pnas.2207987119. Epub 2022 Aug 15. Proc Natl Acad Sci U S A. 2022. PMID: 35969787 Free PMC article.
-
On the difficulties of characterizing weak protein interactions that are critical for neurotransmitter release.FEBS Open Bio. 2022 Nov;12(11):1912-1938. doi: 10.1002/2211-5463.13473. Epub 2022 Sep 2. FEBS Open Bio. 2022. PMID: 35986639 Free PMC article. Review.
-
In Vivo Analysis of a Gain-of-Function Mutation Confirms Unc18/Munc18's Role in Priming.J Neurosci. 2018 Jan 31;38(5):1055-1057. doi: 10.1523/JNEUROSCI.3068-17.2017. J Neurosci. 2018. PMID: 29386300 Free PMC article. Review. No abstract available.
References
-
- Aravamudan B, Fergestad T, Davis WS, Rodesch CK, Broadie K. Drosophila UNC-13 is essential for synaptic transmission. Nat Neurosci. 1999;2:965–971. - PubMed
-
- Ashery U, Betz A, Xu T, Brose N, Rettig J. An efficient method for infection of adrenal chromaffin cells using the Semliki Forest virus gene expression system. Eur J Cell Biol. 1999;78:525–532. - PubMed
-
- Augustin I, Rosenmund C, Südhof TC, Brose N. Munc13-1 is essential for fusion competence of glutamatergic synaptic vesicles. Nature. 1999;400:457–461. - PubMed
-
- Basu J, Shen N, Dulubova I, Lu J, Guan R, Guryev O, Grishin NV, Rosenmund C, Rizo J. A minimal domain responsible for Munc13 activity. Nat Struct Mol Biol. 2005;12:1017–1018. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
