

doi: 10.1038/318635a0.
Primary structure and expression of a functional human glucocorticoid receptor cDNA
- PMID: 2867473
- PMCID: PMC6165583
- DOI: 10.1038/318635a0
Item in Clipboard
Primary structure and expression of a functional human glucocorticoid receptor cDNA
Nature.
.
Abstract
Identification of complementary DNAs encoding the human glucocorticoid receptor predicts two protein forms, of 777 (alpha) and 742 (beta) amino acids, which differ at their carboxy termini. The proteins contain a cysteine/lysine/arginine-rich region which may define the DNA-binding domain. Pure radiolabelled glucocorticoid receptor, synthesized in vitro, is immunoreactive and possesses intrinsic steroid-binding activity characteristic of the native glucocorticoid receptor.
Figures

Human glucocorticoid receptor cDNA sequencing strategy and schematic representation of cDNA clones, a, The composite cDNA for the α glucocorticoid receptor is represented at the top, with noncoding (lines) and coding (stippled portion) sequences indicated. Common 6-nucleotide restriction enzyme sites are shown. Overlapping cDNA inserts used to determine the sequence are shown: arrows beneath the regions sequenced show the direction and extent of sequencing. The dashed line at the 3’ end of OB10 indicates divergent sequence. Numbers refer to nucleotide positions in OB10 relative to the 5’-most transcribed sequence, b, cDNAs encoding the α and β forms of the receptor (OB7 and OB10, respectively). The 5’ end of OB7 (broken lines) is contributed by the OB10 clone. Protein-coding information is represented by wide bars; untranslated sequences are indicated by thin bars. Nucleotides and amino acids are numbered above and below the coding sequence, respectively. Common DNA sequences extend to nucleotide 2,313 (amino-acid residue 727), at which point the α- and β-receptor forms diverge, with the α cDNAs (OB12, OB7) continuing in an open reading frame for 150 nucleotides (50 amino acids) and the β cDNA (OB10) continuing for 45 nucleotides (15 amino acids; see Fig. 3). Hexanucleotide signals (AATAAA) just upstream of the poly(A) in the clones are indicated, with the first hexanucleotide in OB7 serving as poly(A) in OB12 Methods. The inserts hGR1.2, hGR2.9 and hGR5.16 were isolated from a λ gt11 IM-9 lymphoid cell cDNA library as described previously. Two clones were isolated from cDNA libraries constructed by H. Okayama in pcD (ref. 44) using poly(A)+ mRNA from GM637 human fibroblasts (OB7) and primary human fibroblasts (OB10). Screening was performed with the hGR1.2 cDNA, radiolabelled by nick-translation with 32P-dCTP. Sequences were determined by the chemical cleavage method of Maxam and Gilbert.

cDNA and predicted protein sequence of human glucocorticoid receptor. The complete α coding sequence and OB7 3’-untranslated region are shown, with the deduced amino acids given above the long open reading frame. An upstream in-frame stop codon at nucleotides 121–123 and putative additional polyadenylation signals in OB7 are underlined.
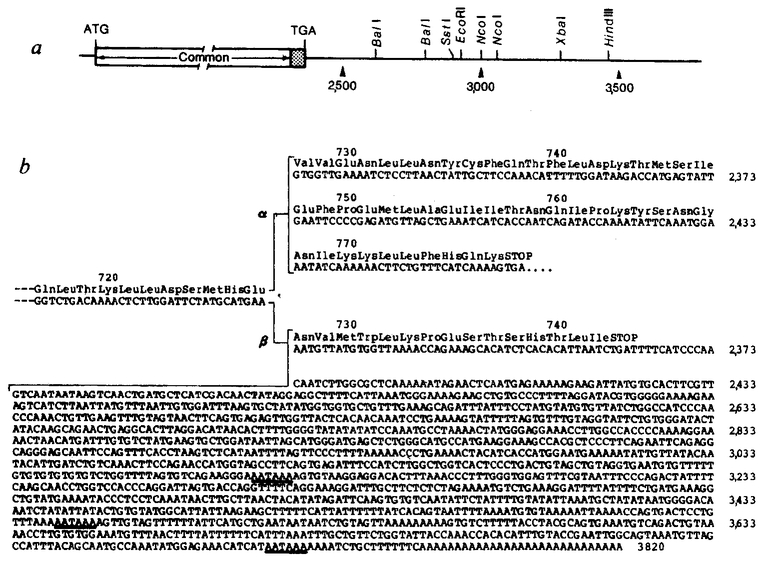
Restriction map and nucleotide sequence of the 3’ end of the human glucocorticoid receptor β cDNA. α, The common 6-nucleotide restriction enzyme sites are shown for the 3’-untranslated region of OB10. b, The cDNA sequence of the β form (OB10) from nucleotide 2,281 to 3,820 compared with the protein-coding information found in the 3’-terminal coding portion of the a form (OB7). Amino acids encoded by each of the cDNAs are presented above the nucleotide sequences. Putative polyadenylation signals (AATAAA) in the 3’-untranslated sequence of OB10 are underlined.

Immunoblot comparison of hGR translated in vitro with in vivo hGR from cell extracts, a, The vectors constructed for in vitro transcription of the hGR cDNA sequence. The complete α (pGR107) and β (pGR108) coding sequences were placed under the transcriptional control of the SP6 promoter in pGEMl. Vector sequences, noncoding cDNA sequences and coding sequences are indicated by thin lines, thick bars and boxed regions, respectively. The poly(A) tract of ~60 nucleotides is indicated by A.,. Divergent coding sequences are indicated by striped and stippled regions, b, Western blot analysis of in vitro translation products and cell extracts. Unlabelled translation products synthesized in a rabbit reticulocyte lysate system with no added RNA (lane 1) or with RNA synthesized from pGR108 (β, lane 2) or pGR107 (α, lane 3) were fractionated on a 7.5% SDS-polyacrylamide gel. Additional lanes are: cytoplasmic extracts from IM-9 (lane 4), IM-9 treated with 1 μM triamcinolone acetonide (lane 5), HeLa (lane 6), ADR6.M1890.AD1 mouse lymphoma (lane 7), S49 mouse lymphoma (lane 8) and EL4 lymphoma (lane 9). Proteins were transferred to nitrocellulose and probed with anti-hGR antibody, followed by 125I-labelIed Staphylococcus aureus protein A as described previously. Methods. To construct an expression vector containing the entire α coding sequence shown in Fig. 2, the 3’ coding sequence of OB7 was fused to OB10 5’ coding information. OB7 was partially digested with EcoRI, completely digested with X baI, and the 1.2-kbp fragment was gel-purified and ligated with EcoRI/baI-digested OB10 to produce the intermediate pOB107. The entire pOBl07 cDNA sequence including the 5’ poly(G) tract (11 nucleotides, nt) and 3’ poly(A) tract (~ 6 0 nt) was excised by partial Pst I/complete Bam HI digestion. The resultant 3.5-kb fragment was gel-purified and inserted between the PstI and Bam HI sites of pGEM l (Promega Biotec) to yield pGR107. Plasmid GR108 was directly constructed from pOB10 by partial PstI/com plete Bam HI digestion and insertion of the resulting cDNA insert into the corresponding sites of pGEMl. Capped SP6 transcripts were synthesized from PvuII-linearized pGR107 and pGR108, as described by Kjieg and Melton, with simultaneous capping effected by reduction of the GTP concentration from 400 to 100 μM and addition of m7GppG (Pharmacia) to 500 μM. Transcripts were purified by P60 chromatography and translated with micrococcal nuclease-treated rabbit reticulocyte lysate (Promega Biotec) in conditions suggested by the manufacturer. Preparation of IM-9 cytosol from steroid-treated cells was as described previously. Size markers are phosphorylase B (97K), bovine serum albumin (66K.) and ovalbumin (45K).

Steroid-binding of α-hGR (GR107) translated in vitro. Binding to IM-9 cytosol extract (stippled bars) and to reticulocyte lysate containing SP6-generated α-hGR RNA (GR107; open bars) are shown. Bars represent bound 3H-triamcinolone acetonide (TA) determined with a 100-fold excess of various steroid competitors; 100% competition was determined using unlabelled TA as competitor. The values represent the mean of triplicate determinations, with error bars showing P
< 0.05. Steroid competitors are dexamethasone (Dex), cortisol (Cort), progesterone (Prog), testosterone (Test), and oestradiol (Oest). Methods. Binding assays were performed in 100 μl containing 10 mM Tris-HCl pH 7.4, 100 mM NaCl, 1 mM EDTA, 10 mM sodium molybdate, 10 dithiothreitol, 150 mM 3H-TA (20 Ci mmol−1; Amersham) and 10 μl translation mixture or 100 μg fresh IM-9 cytosol. Unlabelled steroid competitor (15 μM ) was added as indicated. After 2 h at 0 °C, samples were extracted twice for 5 min each with 5 μl of 50% dextran-coated charcoal to remove unbound steroid, and counted. Uncompeted and fully competed values for the α glucocorticoid receptor (GR107) were 490 and 290 c.p.m., respectively. Reticulocyte lysate translation mixtures without added transcript or programmed with β-receptor SP6 RNA (GR108) contained no competable 3H-TA binding.

Chromosome mapping analysis of hGR cDNA. A, 10 μg of DNA from human placenta (lanes 1, 4), CHO/human somatic cell hybrid (HHW454, lanes 2, 5) containing chromosome 5 as its only human complement, or CHO (lanes 3, 6), was digested with EcoRI (lanes 1–3) or HindIII (lanes 4–6) to completion, fractionated on a 0.8% agarose gel and transferred to nitrocellulose paper. b, Chromosomes (3 × 104) prepared from a human lymphocyte cell line, stained with 4,6-bis(2”-imadazolinyl-4H,5H)-2-phenylindole (DIPI)/chromomycin A3 and sorted using a dual-laser custom FACS IV chromosome sorter, were denatured and neutralized on nitrocellulose paper. Note that Hoechst/chromomycin-stained chromosome 9 was sorted with chromosomes 10–12. c, 10 μg of DNA from the parental mouse cell line MEL (lane 1) or the parentally derived somatic cell hybrid carrying human chromosome 16 (ref. ; lane 2) was digested with HindIII and separated on a 0.8% agarose gel then transferred to nitrocellulose paper. All filters were probed with the 1,100-bp insert from hGR 1.2, nick-translated to a specific activity of 3 × 108 c.p.m.μg−l and hybridized in 5 × SSPE, 1 × Denhardt’s, 0.1% SDS, 50% formamide, 100 μg ml−1 denatured salmon sperm DNA, 50% dextran sulphate at 42 °C for 18 h. Filters were washed twice (for 30 min each) in 2 × SSC at 68 °C and exposed to X-ray film at −70 °C with an intensifying screen.
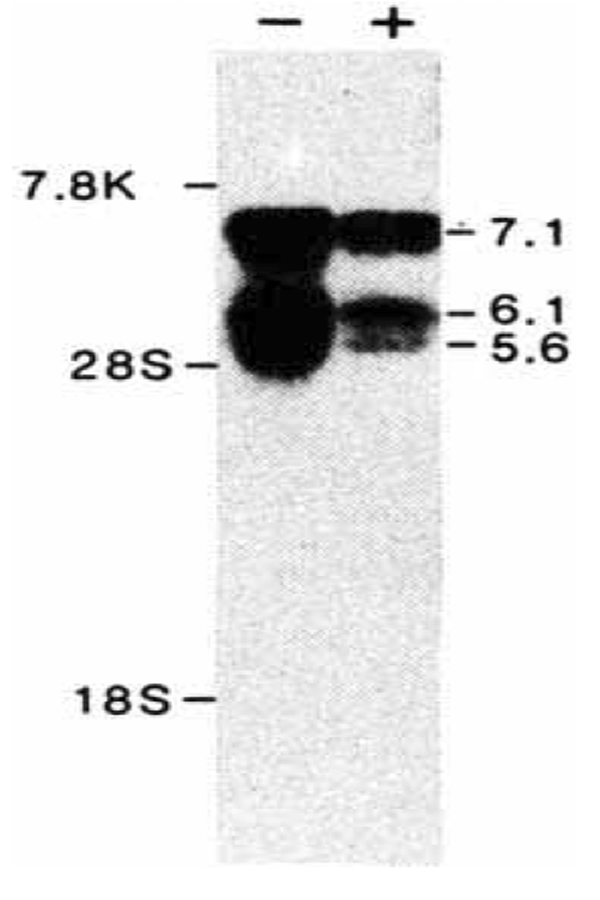
Northern blot analysis of hGR mRNA. 10 μg of poly(A)+ mRNA from human HT1080 fibroblast cells, collected after 24 h without ( − ) or with ( + ) treatment with 10 μM dexamethasone, was electrophoresed through a 0.8% agarose/1% formaldehyde gel, stained with acridine orange (120 μg ml−1) and transferred to nitrocellulose. The filter was hybridized overnight with nick-translated hGR1.2 (106 c.p.m. ml−1, specific activity 108 c.p.m. μg−1) and washed with 2 × SSC at 68 °C. Sizes were estimated from human fibronectin mRNA (7.8 kb), and 28 S (5.0 kb) and 18 S (2.1 kb) ribosomal RNAs.
Similar articles
-
Domain structure of human glucocorticoid receptor and its relationship to the v-erb-A oncogene product.Nature. 1985 Dec 19-1986 Jan 1;318(6047):670-2. doi: 10.1038/318670a0. Nature. 1985. PMID: 3841189
-
Human steroid receptors and erb-A gene products form a superfamily of enhancer-binding proteins.Clin Physiol Biochem. 1987;5(3-4):179-89. Clin Physiol Biochem. 1987. PMID: 3304776
-
Intragenic sequences of the human glucocorticoid receptor complementary DNA mediate hormone-inducible receptor messenger RNA down-regulation through multiple mechanisms.Mol Endocrinol. 1994 Dec;8(12):1764-73. doi: 10.1210/mend.8.12.7708063. Mol Endocrinol. 1994. PMID: 7708063
-
DNA-binding by the glucocorticoid receptor: a structural and functional analysis.J Steroid Biochem Mol Biol. 1992 Mar;41(3-8):249-72. doi: 10.1016/0960-0760(92)90351-i. J Steroid Biochem Mol Biol. 1992. PMID: 1562506 Review.
-
The structure of the human glucocorticoid receptor and its gene.J Steroid Biochem. 1987;27(1-3):105-8. doi: 10.1016/0022-4731(87)90300-1. J Steroid Biochem. 1987. PMID: 2447387 Review.
Cited by
-
Early onset pre-eclampsia is associated with altered DNA methylation of cortisol-signalling and steroidogenic genes in the placenta.PLoS One. 2013 May 7;8(5):e62969. doi: 10.1371/journal.pone.0062969. Print 2013. PLoS One. 2013. PMID: 23667551 Free PMC article.
-
Signaling by nuclear receptors.Cold Spring Harb Perspect Biol. 2013 Mar 1;5(3):a016709. doi: 10.1101/cshperspect.a016709. Cold Spring Harb Perspect Biol. 2013. PMID: 23457262 Free PMC article. Review.
-
Obestatin signalling counteracts glucocorticoid-induced skeletal muscle atrophy via NEDD4/KLF15 axis.J Cachexia Sarcopenia Muscle. 2021 Apr;12(2):493-505. doi: 10.1002/jcsm.12677. Epub 2021 Mar 9. J Cachexia Sarcopenia Muscle. 2021. PMID: 33687156 Free PMC article.
-
Deletion mapping of DNA markers to a region of chromosome 5 that cosegregates with schizophrenia.Genomics. 1989 Nov;5(4):940-4. doi: 10.1016/0888-7543(89)90138-9. Genomics. 1989. PMID: 2591972 Free PMC article.
-
Differential regulation by dexamethasone of glucocorticoid receptor messenger RNA concentrations in neuronal cultures derived from fetal rat hypothalamus and cerebral cortex.Cell Mol Neurobiol. 1990 Jun;10(2):227-35. doi: 10.1007/BF00734576. Cell Mol Neurobiol. 1990. PMID: 2364410
References
-
- Jensen EV & De Sombre ER A. Rev. Biochem 41, 203–230 (1972). - PubMed
-
- Gorski J & Gannon FA, Rev. Physiol 38, 425–450 (1976). - PubMed
-
- Yamamoto K,R & Alberts BM A. Rev. Biochem 45, 721–746 (1976). - PubMed
-
- O’Malley BW, McGuire WL, Kohler PO & Komtman SG Recent Prog. Horm. Res 25, 105–160 (1969). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
