

doi: 10.1007/s12551-014-0150-6.
Epub 2015 Feb 13.
Kinesin Motor Enzymology: Chemistry, Structure, and Physics of Nanoscale Molecular Machines
Affiliations
- PMID: 28510227
- PMCID: PMC5418420
- DOI: 10.1007/s12551-014-0150-6
Item in Clipboard
Kinesin Motor Enzymology: Chemistry, Structure, and Physics of Nanoscale Molecular Machines
Biophys Rev.
2015 Sep.
Abstract
Molecular motors are enzymes that convert chemical potential energy into controlled kinetic energy for mechanical work inside cells. Understanding the biophysics of these motors is essential for appreciating life as well as apprehending diseases that arise from motor malfunction. This review focuses on kinesin motor enzymology with special emphasis on the literature that reports the chemistry, structure and physics of several different kinesin superfamily members.
Keywords: Chemical models; Enzymes; Kinesin motor enzymology; Kinesin superfamily; Molecular motors.
Figures
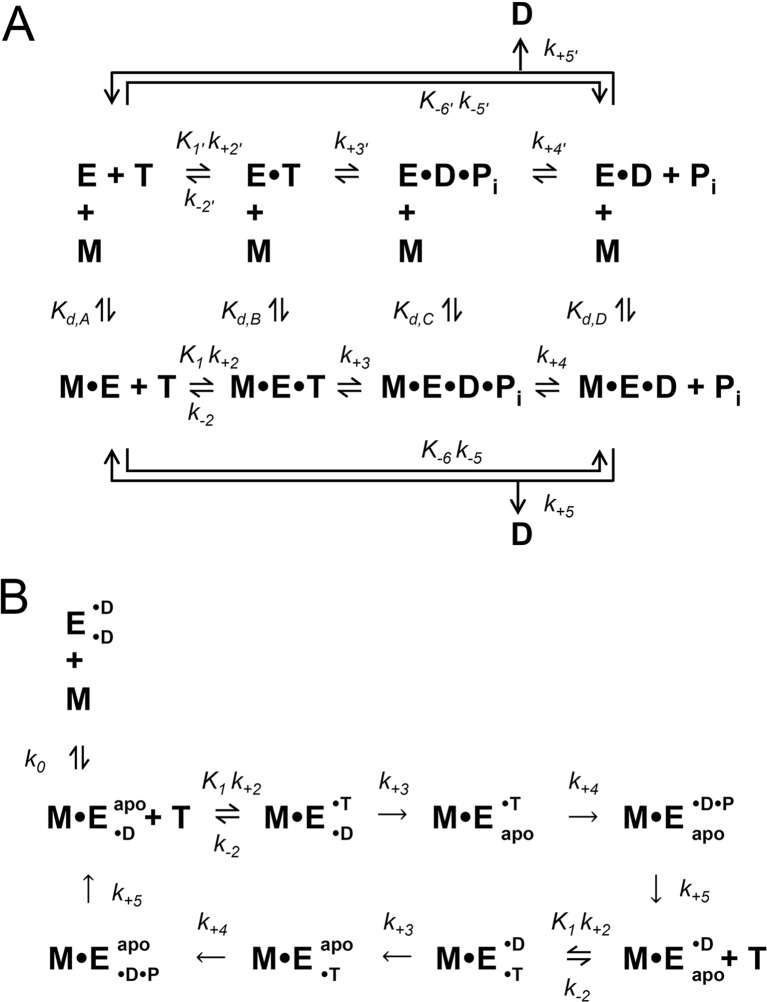
Minimal ATPase mechanisms for monomeric and dimeric kinesins. Monomeric (a) and dimeric (b) kinesin ATPase mechanisms are shown. E kinesin, M microtubule, T ATP substrate, D ADP product, P
i inorganic phosphate product, apo nucleotide-free/rigor. For dimeric kinesins that have two inter-molecularly regulated active sites, each site is depicted as either a superscript or subscript (for example: E• ADP• ADP indicates tha3 both sites of the dimeric kinesin (E) are bound to ADP)
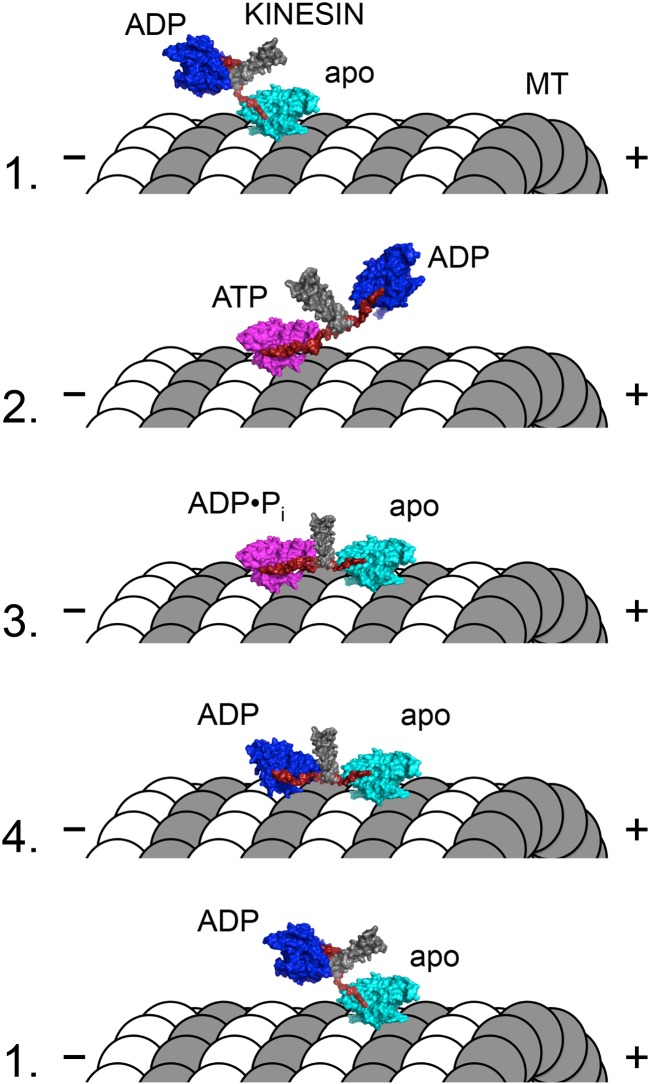
Model for the kinesin-1 “hand-over-hand” walking mechanism. Molecular representations of the dimeric kinesin-1 motor and microtubule (MT) are shown in each of four states (labeled numbers to the left of figure). The coiled–coil stalk (gray) and neck linker (red) are highlighted in each state. Coloring of the kinesin motor core represents three structural states: dark blue strongly ADP-bound, weakly microtubule-bound, neck linker undocked; cyan weakly nucleotide-bound, strongly microtubule-bound, neck linker undocked; magenta strongly ATP- or ADP•Pi-bound, strongly microtubule-bound, neck linker docked. Dimeric kinesin-1 motors are purified with ADP tightly bound to both active sites. The first head to strongly interact with the microtubule (head 1) will rapidly release its bound ADP to reach the nucleotide-free/apo/rigor state (state 1). Since head 2 remains unattached to the microtubule, head 1 is able to bind ATP, which docks the neck linker of head 1, biasing head 2 forward (i.e., toward the MT plus-end) to the next binding site on the same MT protofilament (state 2). ATP hydrolysis on head 1 is concomitant with tight microtubule-binding by head 2 to rapidly release its ADP product (state 3). Upon Pi release from head 1, intramolecular strain coupled to conformational changes in the ADP state of head 1 lead to weakening of head 1’s interaction with the MT (state 3). After detachment of head 1 from the MT, the kinesin effectively reaches state 1 with the exception that the heads trade places (i.e., head 2 is nucleotide-free/apo/rigor while head 1 is ADP-bound and detached from the MT). The transition from state 3 to state 4 leads to a release of the intramolecular tension that is thought to provide a mechanical gating of ATP binding to the front head
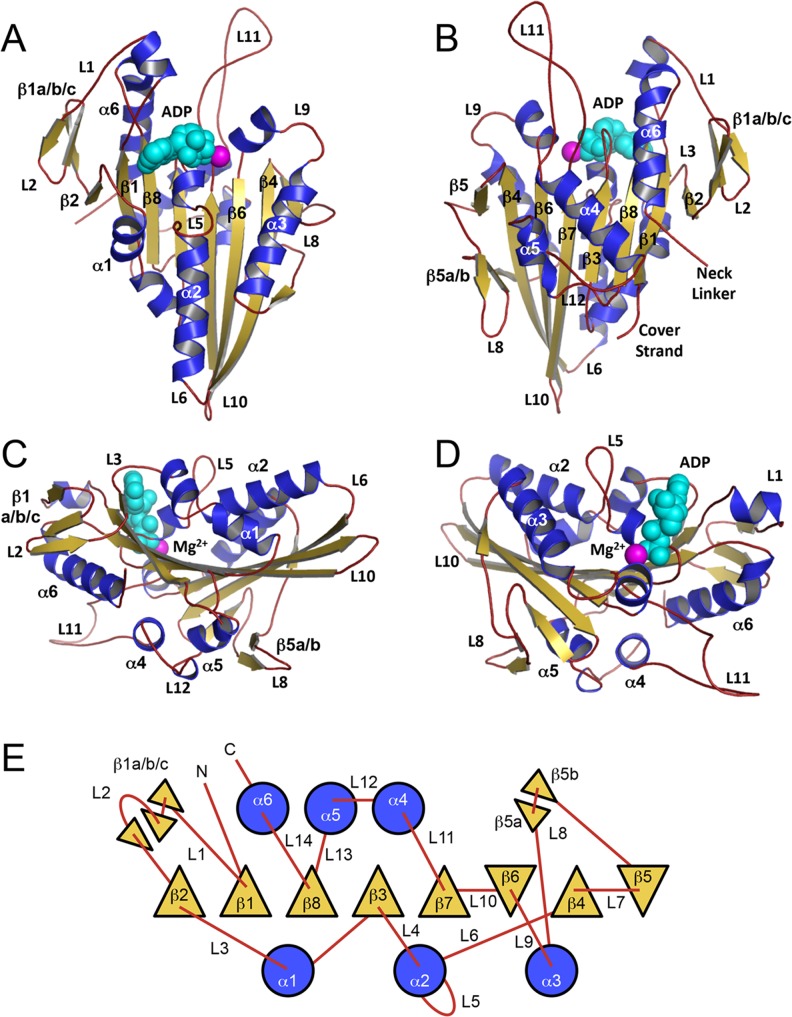
Kinesin structure and topology. The monomeric human kinesin-1 structure (1BG2; Kull et al. 1996) is depicted in four distinct orientations: a front, b back, c left side, d right side. Secondary structure elements (α-helices, β-strands, and loops) are colored blue, gold, and red, respectively. Mg2+ and ADP are shown as space-fill model and are colored magenta and cyan, respectively. e Topology diagram of the kinesin motor domain shows α-helices (circles; shaded blue; up/down direction indicated by loop attachment), β-strands (triangles; shaded gold; up/down to depict orientation), and loops (red lines) as rendered from (Kull et al. 1998)
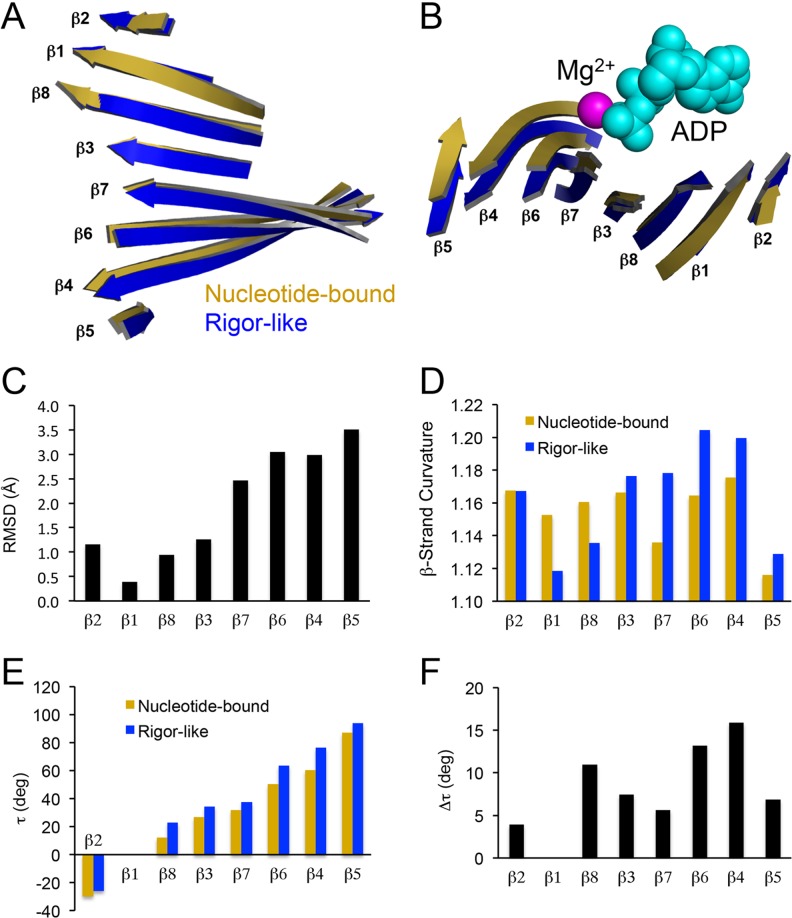
Structural changes in the eight-stranded central β-sheet of kinesin. Side (a) and end (b) views of the kinesin β-sheet after a nucleotide-bound structure (1BG2; Kull et al. 1996) was aligned with a rigor-like structure (4OZQ; Arora et al. 2014) using the main chain atoms of the P-loop. Mg2+ and ADP are shown as space-fill model and are colored magenta and cyan, respectively. c The root-mean-square deviation (RMSD) of complementary residues is plotted for each individual β-strand. d Individual β-strand curvature based on the sum of sequential Cα distances for each β-strand residue (i = initial residue, i + x = final residue) divided by the total distance from initial to final plotted for nucleotide-bound and rigor-like structures. e β-strand torsion angles (τ) relative to β1 are plotted to quantify the degree of twisting within the entire β-sheet. Torsion angles were calculated using the following equation for each individual β-strand: , where are the unit vectors for each respective β-strand (Cecchini et al. 2008). f The change in each individual β-strand torsion angle (Δτ) as the kinesin central β-sheet transitions from nucleotide-bound (1BG2) to rigor-like (4OZQ) states
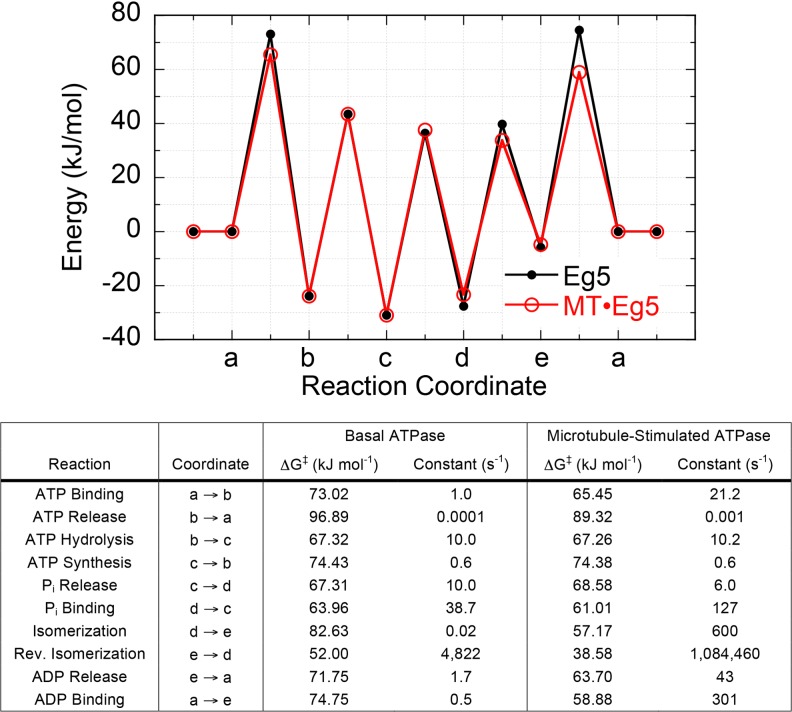
Kinesin energy landscape in the absence and presence of microtubules. The energy landscapes for human monomeric kinesin-5 (Eg5) in the absence (black) and presence (red) of microtubules are shown. The thermodynamically and kinetically defined intermediate states are defined in the table. The magnitude of energy for each transition state was calculated from the Arrhenius/Eyring/Kramer equation using previously determined kinetic rate constants for the monomeric kinesin-5 ATPase mechanism (Table 1). Adapted and updated from Zhao et al. (2010)
Similar articles
-
Kinesin motors as molecular machines.Bioessays. 2003 Dec;25(12):1212-9. doi: 10.1002/bies.10358. Bioessays. 2003. PMID: 14635256 Review.
-
Molecular Assembly of Rotary and Linear Motor Proteins.Acc Chem Res. 2019 Jun 18;52(6):1623-1631. doi: 10.1021/acs.accounts.9b00015. Epub 2019 Mar 18. Acc Chem Res. 2019. PMID: 30882207
-
Collective dynamics of kinesin.Phys Rev E Stat Nonlin Soft Matter Phys. 2009 Mar;79(3 Pt 1):031929. doi: 10.1103/PhysRevE.79.031929. Epub 2009 Mar 31. Phys Rev E Stat Nonlin Soft Matter Phys. 2009. PMID: 19391993
-
Subunits interactions in kinesin motors.Eur J Cell Biol. 2007 Sep;86(9):559-68. doi: 10.1016/j.ejcb.2007.05.008. Epub 2007 Jul 12. Eur J Cell Biol. 2007. PMID: 17628208 Review.
-
Experimental and theoretical energetics of walking molecular motors under fluctuating environments.Biophys Rev. 2020 Apr;12(2):503-510. doi: 10.1007/s12551-020-00684-7. Epub 2020 Mar 16. Biophys Rev. 2020. PMID: 32173796 Free PMC article. Review.
Cited by
-
Shaft Function of Kinesin-1's α4 Helix in the Processive Movement.Cell Mol Bioeng. 2019 Jun 25;12(4):345-354. doi: 10.1007/s12195-019-00581-4. eCollection 2019 Aug. Cell Mol Bioeng. 2019. PMID: 31719918 Free PMC article.
-
Sequences in the stalk domain regulate auto-inhibition and ciliary tip localization of the immotile kinesin-4 KIF7.J Cell Sci. 2021 Jul 1;134(13):jcs258464. doi: 10.1242/jcs.258464. Epub 2021 Jul 8. J Cell Sci. 2021. PMID: 34114033 Free PMC article.
-
Comparison of explicit and mean-field models of cytoskeletal filaments with crosslinking motors.Eur Phys J E Soft Matter. 2021 Mar 29;44(3):45. doi: 10.1140/epje/s10189-021-00042-9. Eur Phys J E Soft Matter. 2021. PMID: 33779863 Free PMC article.
-
How Kinesin-1 Utilize the Energy of Nucleotide: The Conformational Changes and Mechanochemical Coupling in the Unidirectional Motion of Kinesin-1.Int J Mol Sci. 2020 Sep 22;21(18):6977. doi: 10.3390/ijms21186977. Int J Mol Sci. 2020. PMID: 32972035 Free PMC article. Review.
-
New insights into the mechanochemical coupling mechanism of kinesin-microtubule complexes from their high-resolution structures.Biochem Soc Trans. 2023 Aug 31;51(4):1505-1520. doi: 10.1042/BST20221238. Biochem Soc Trans. 2023. PMID: 37560910 Free PMC article. Review.
References
-
- Admiraal SJ, Herschlag D. Mapping the transition state for ATP hydrolysis: implications for enzymatic catalysis. Chem Biol. 1995;2:729–739. - PubMed
-
- Arnal I, Metoz F, DeBonis S, Wade RH. Three-dimensional structure of functional motor proteins on microtubules. Curr Biol. 1996;6:1265–1270. - PubMed
-
- Asenjo AB, Chatterjee C, Tan D, et al (2013) Structural model for tubulin recognition and deformation by kinesin-13 microtubule depolymerases. Cell Rep 3:759–768. doi:10.1016/j.celrep.2013.01.030 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
