

DUX-family transcription factors regulate zygotic genome activation in placental mammals
- PMID: 28459456
- PMCID: PMC5446900
- DOI: 10.1038/ng.3858
DUX-family transcription factors regulate zygotic genome activation in placental mammals
Abstract
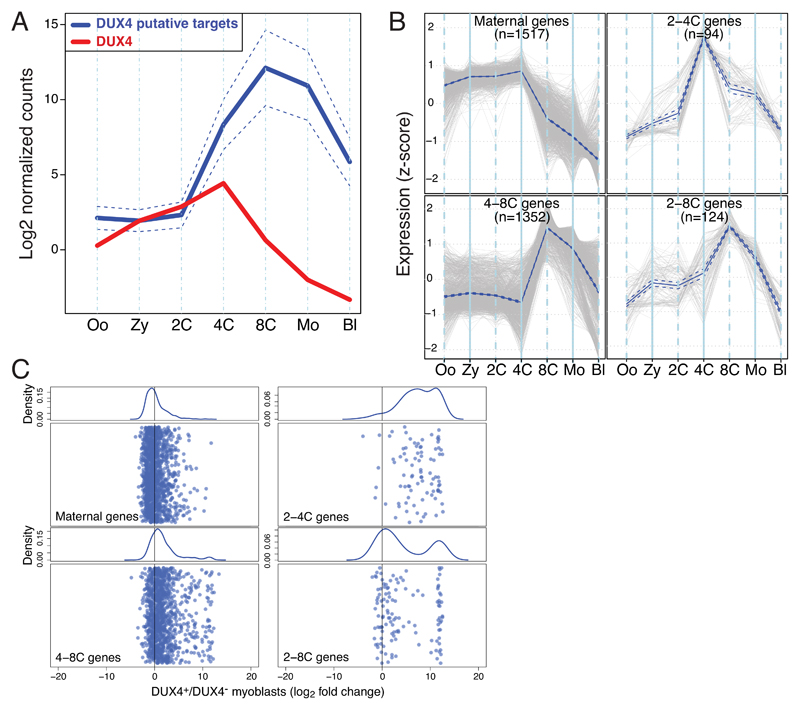
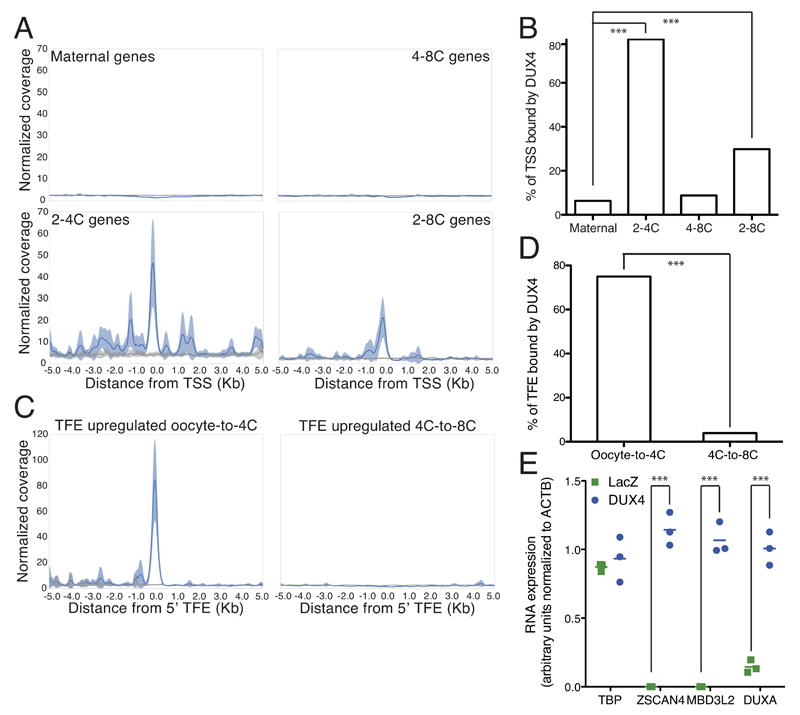
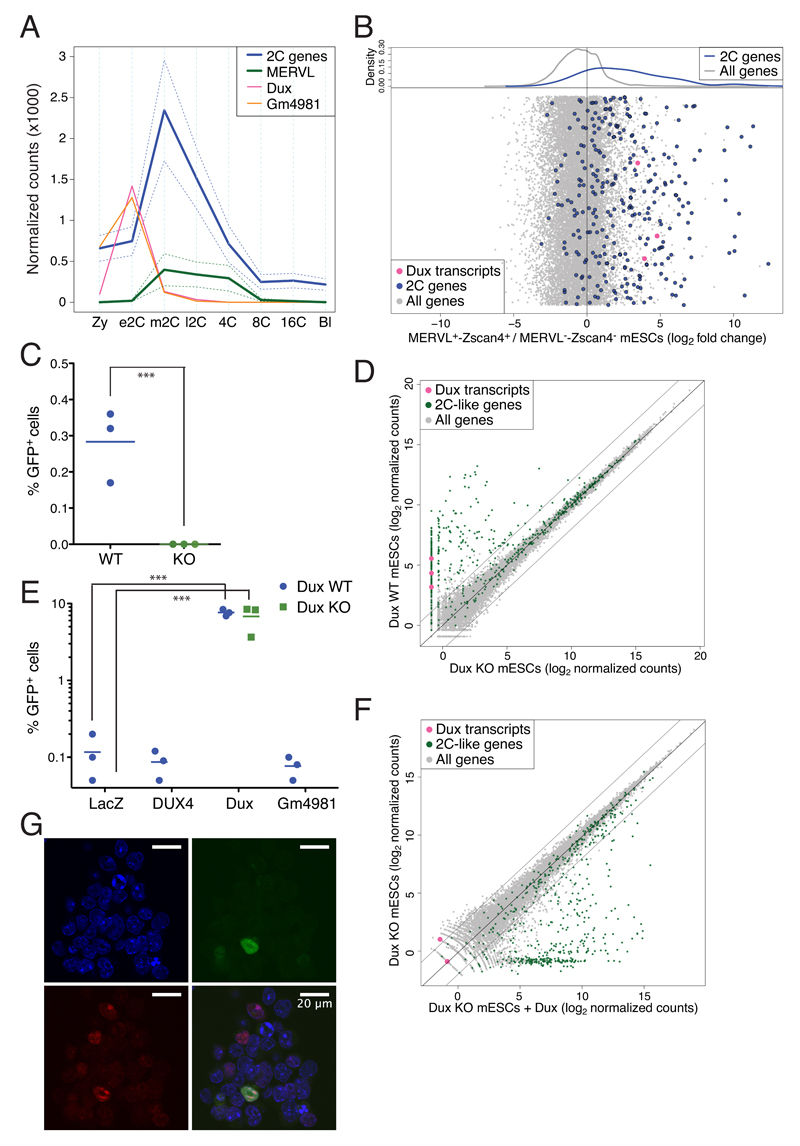
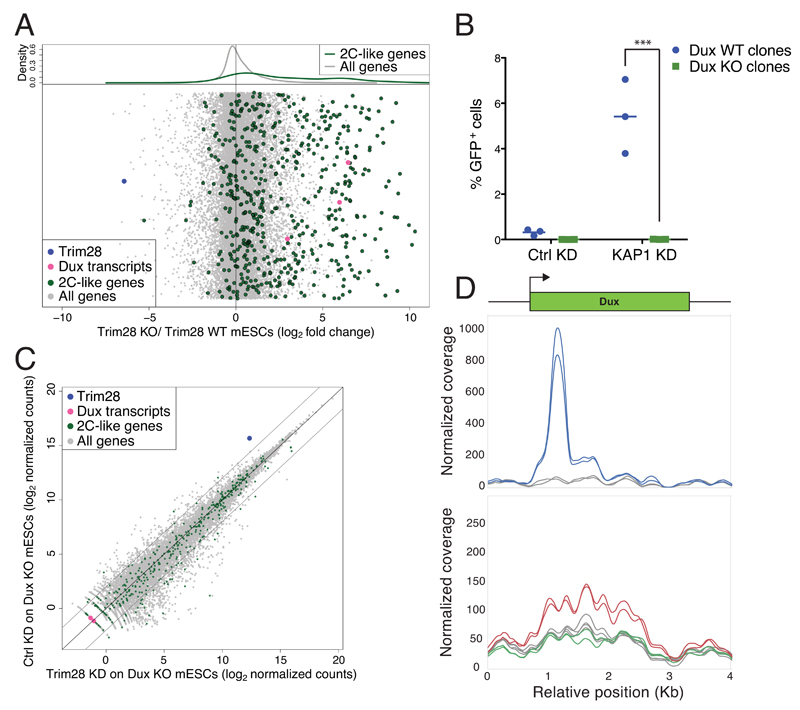
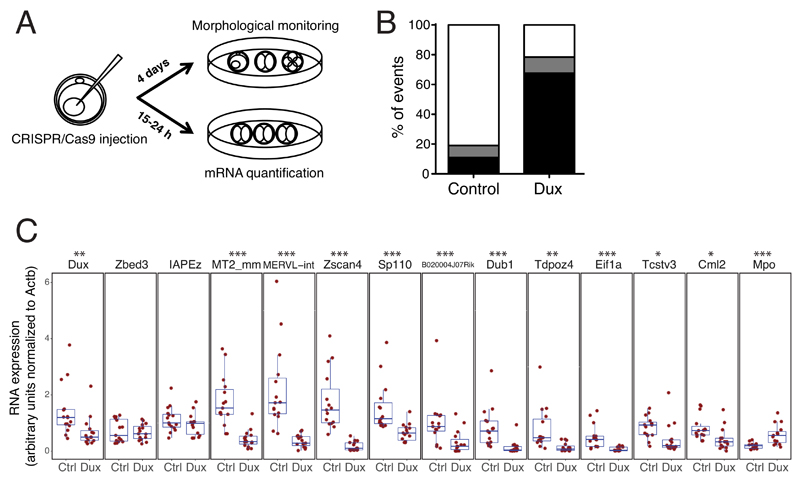
In animal embryos, transcription is mostly silent for several cell divisions, until the release of the first major wave of embryonic transcripts through so-called zygotic genome activation (ZGA). Maternally provided ZGA-triggering factors have been identified in Drosophila melanogaster and Danio rerio, but their mammalian homologs are still undefined. Here, we provide evidence that the DUX family of transcription factors is essential to this process in mice and potentially in humans. First, human DUX4 and mouse Dux are both expressed before ZGA in their respective species. Second, both orthologous proteins bind the promoters of ZGA-associated genes and activate their transcription. Third, Dux knockout in mouse embryonic stem cells (mESCs) prevents the cells from cycling through a 2-cell-like state. Finally, zygotic depletion of Dux leads to impaired early embryonic development and defective ZGA. We conclude that DUX-family proteins are key inducers of zygotic genome activation in placental mammals.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Dppa2 and Dppa4 directly regulate the Dux-driven zygotic transcriptional program.Genes Dev. 2019 Feb 1;33(3-4):194-208. doi: 10.1101/gad.321174.118. Epub 2019 Jan 28. Genes Dev. 2019. PMID: 30692203 Free PMC article.
-
Loss of DUX causes minor defects in zygotic genome activation and is compatible with mouse development.Nat Genet. 2019 Jun;51(6):947-951. doi: 10.1038/s41588-019-0418-7. Epub 2019 May 27. Nat Genet. 2019. PMID: 31133747 Free PMC article.
-
p53 convergently activates Dux/DUX4 in embryonic stem cells and in facioscapulohumeral muscular dystrophy cell models.Nat Genet. 2021 Aug;53(8):1207-1220. doi: 10.1038/s41588-021-00893-0. Epub 2021 Jul 15. Nat Genet. 2021. PMID: 34267371 Free PMC article.
-
DUX: One Transcription Factor Controls 2-Cell-like Fate.Int J Mol Sci. 2022 Feb 13;23(4):2067. doi: 10.3390/ijms23042067. Int J Mol Sci. 2022. PMID: 35216182 Free PMC article. Review.
-
Pioneer Transcription Factors: The First Domino in Zygotic Genome Activation.Biomolecules. 2024 Jun 18;14(6):720. doi: 10.3390/biom14060720. Biomolecules. 2024. PMID: 38927123 Free PMC article. Review.
Cited by
-
Endogenous retroviruses shape pluripotency specification in mouse embryos.Sci Adv. 2024 Jan 26;10(4):eadk9394. doi: 10.1126/sciadv.adk9394. Epub 2024 Jan 24. Sci Adv. 2024. PMID: 38266080 Free PMC article.
-
Transplantation of PSC-derived myogenic progenitors counteracts disease phenotypes in FSHD mice.NPJ Regen Med. 2022 Sep 2;7(1):43. doi: 10.1038/s41536-022-00249-0. NPJ Regen Med. 2022. PMID: 36056021 Free PMC article.
-
The prospects of targeting DUX4 in facioscapulohumeral muscular dystrophy.Curr Opin Neurol. 2020 Oct;33(5):635-640. doi: 10.1097/WCO.0000000000000849. Curr Opin Neurol. 2020. PMID: 32796277 Free PMC article. Review.
-
Crystal Structure of the Double Homeodomain of DUX4 in Complex with DNA.Cell Rep. 2018 Dec 11;25(11):2955-2962.e3. doi: 10.1016/j.celrep.2018.11.060. Cell Rep. 2018. PMID: 30540931 Free PMC article.
-
Gene Editing to Tackle Facioscapulohumeral Muscular Dystrophy.Front Genome Ed. 2022 Jul 15;4:937879. doi: 10.3389/fgeed.2022.937879. eCollection 2022. Front Genome Ed. 2022. PMID: 35910413 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
