

Polyglutamine tracts regulate beclin 1-dependent autophagy
- PMID: 28445460
- PMCID: PMC5420314
- DOI: 10.1038/nature22078
Polyglutamine tracts regulate beclin 1-dependent autophagy
Abstract
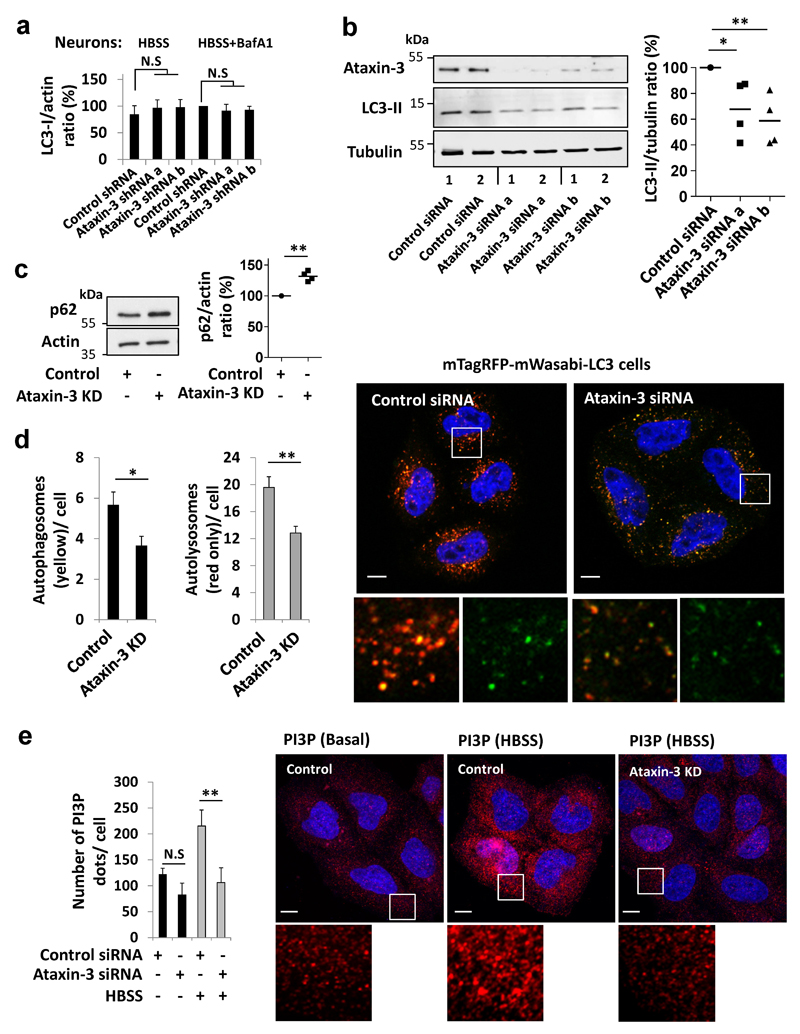
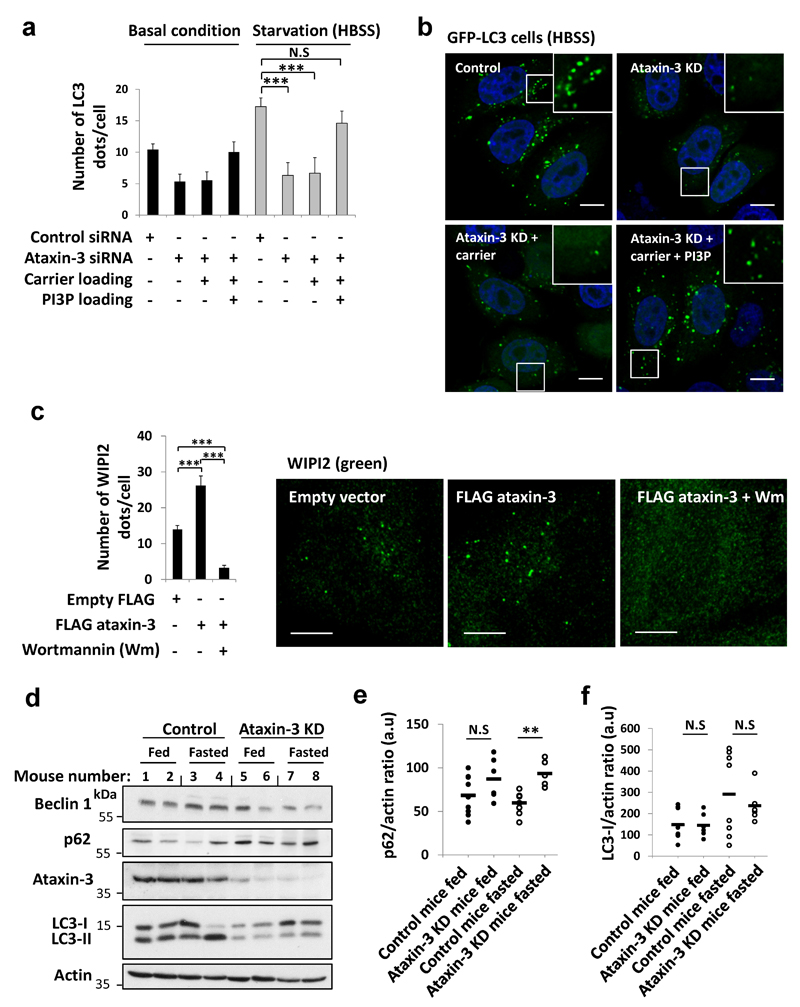
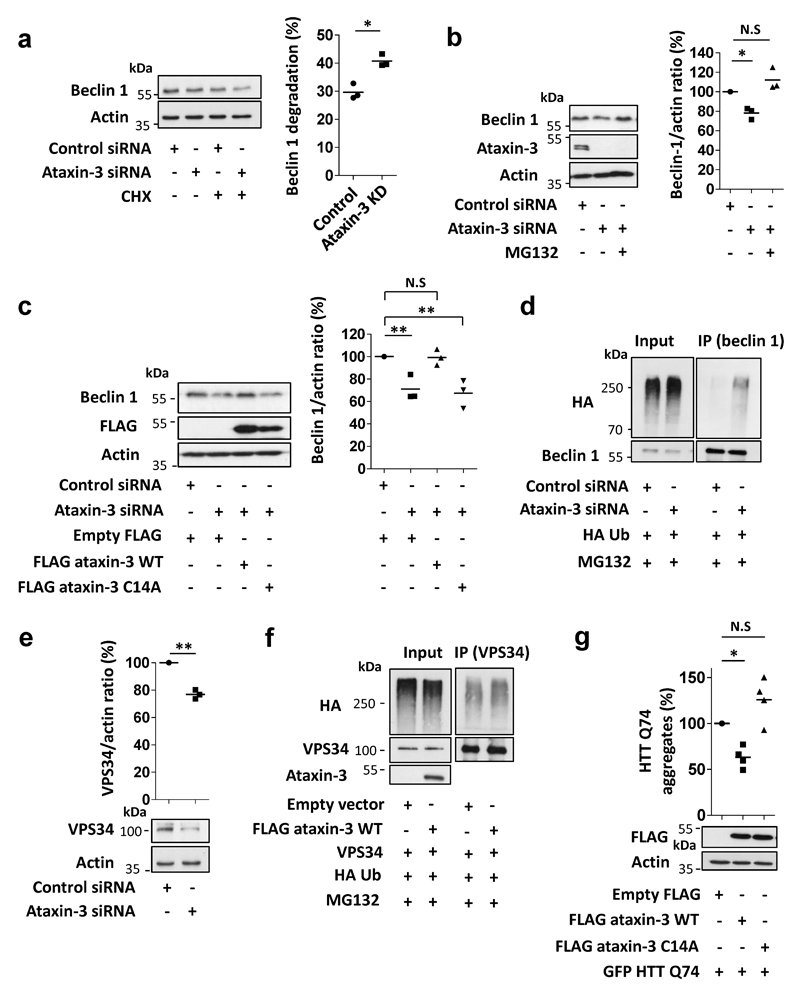
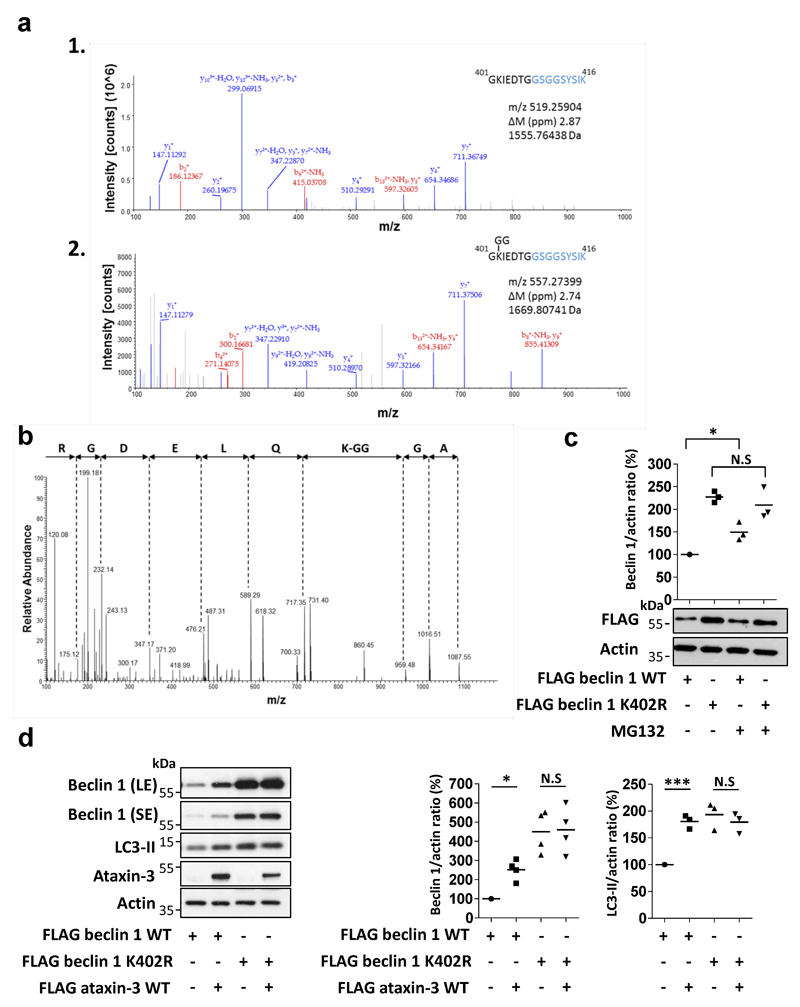
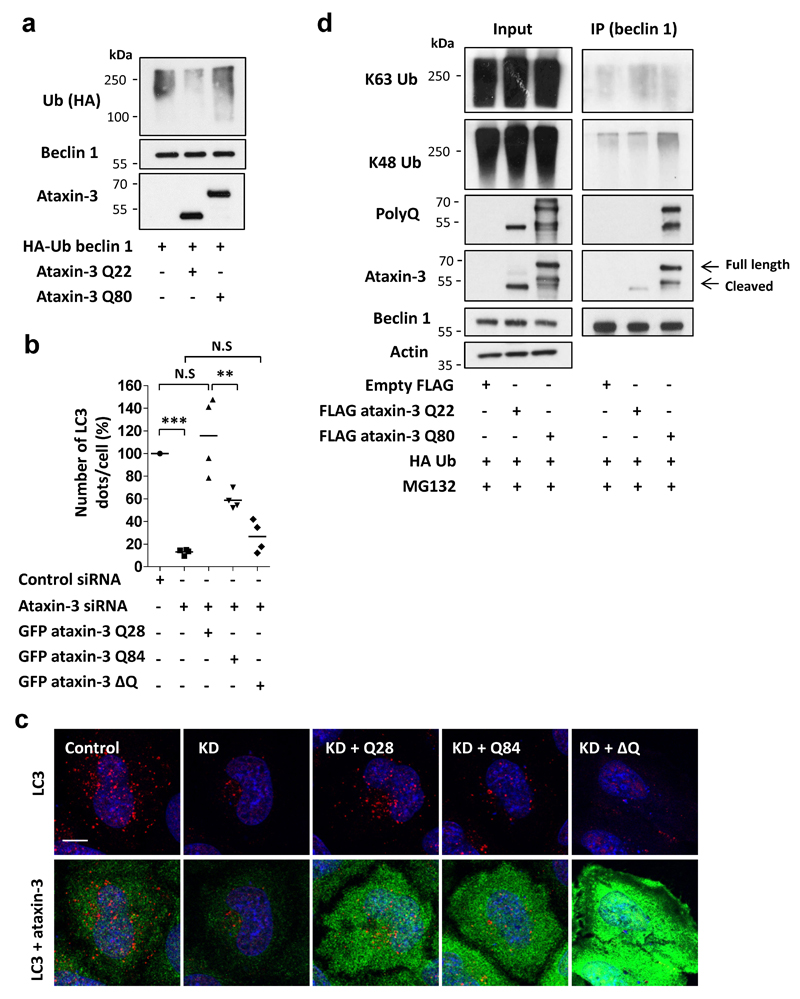
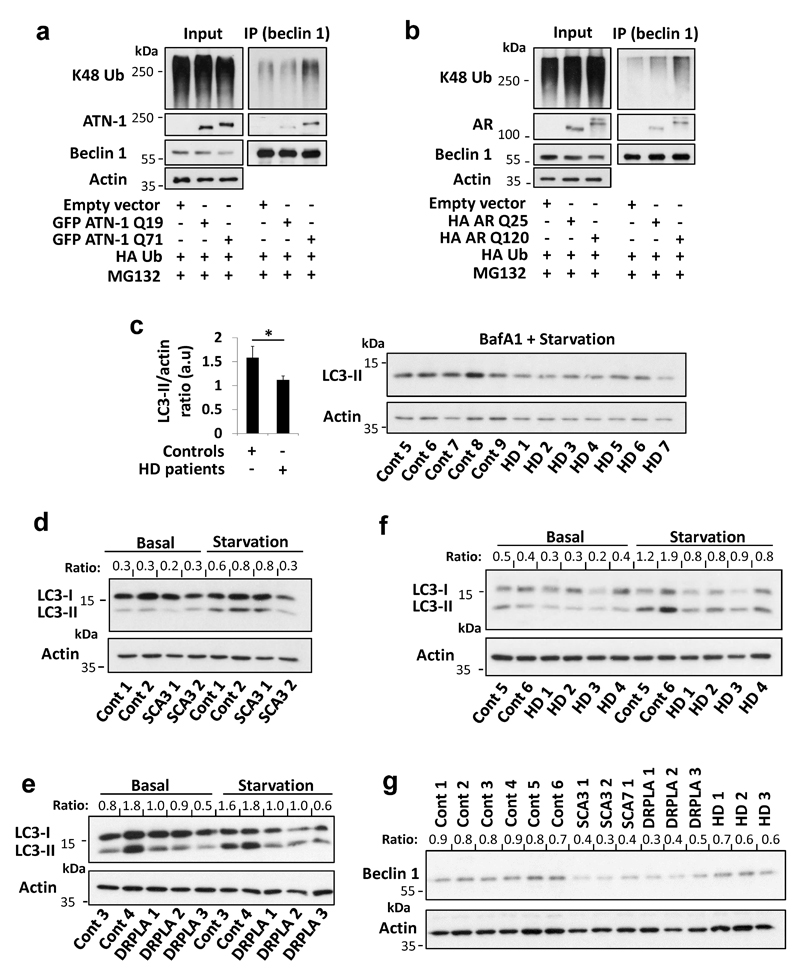
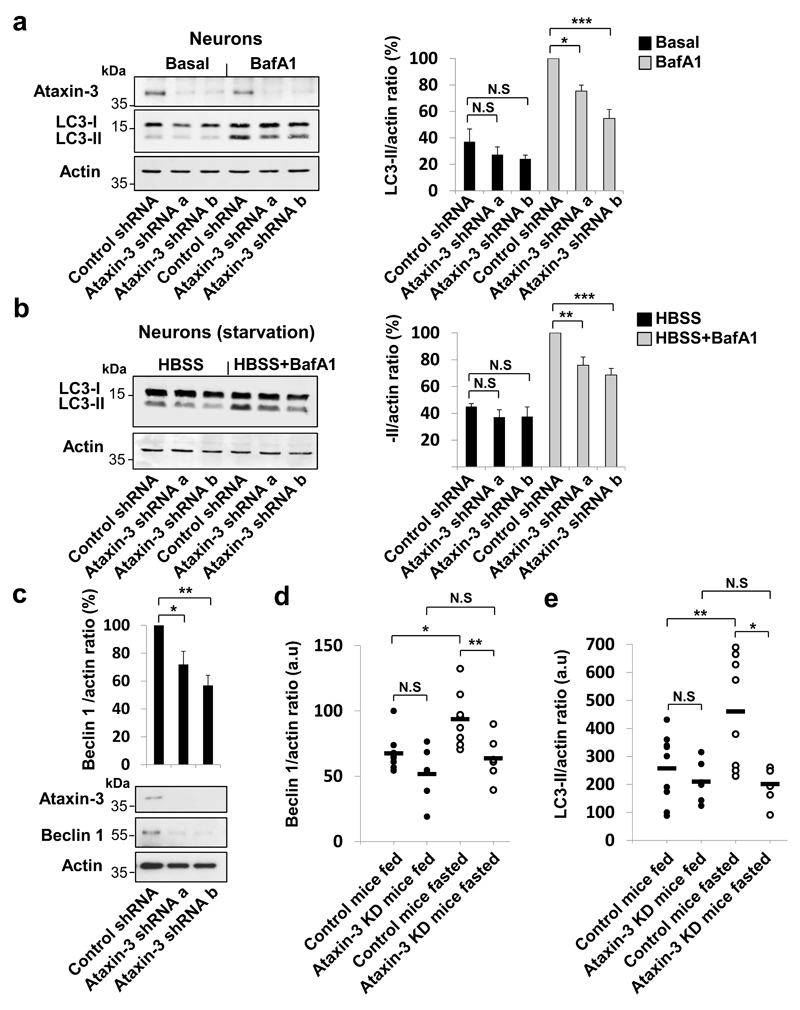
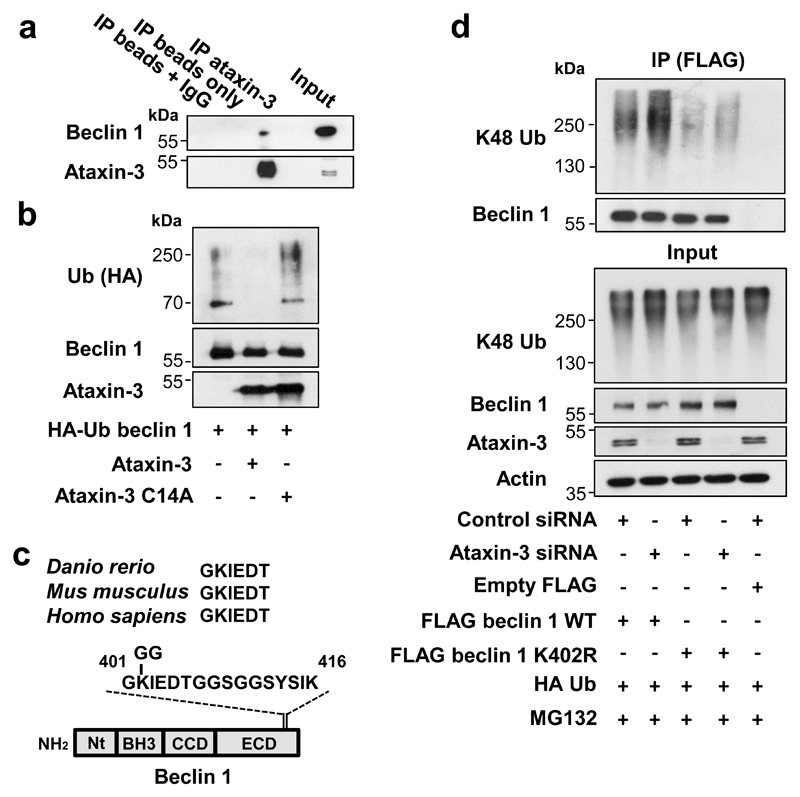
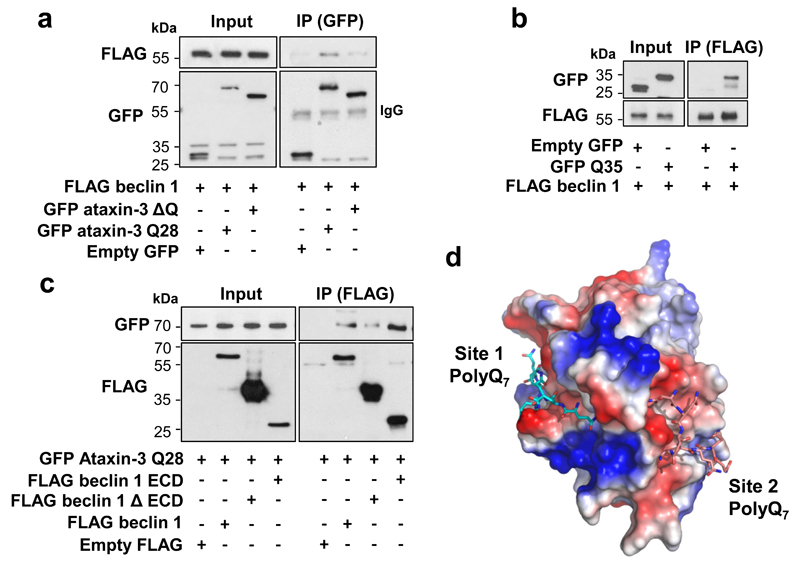
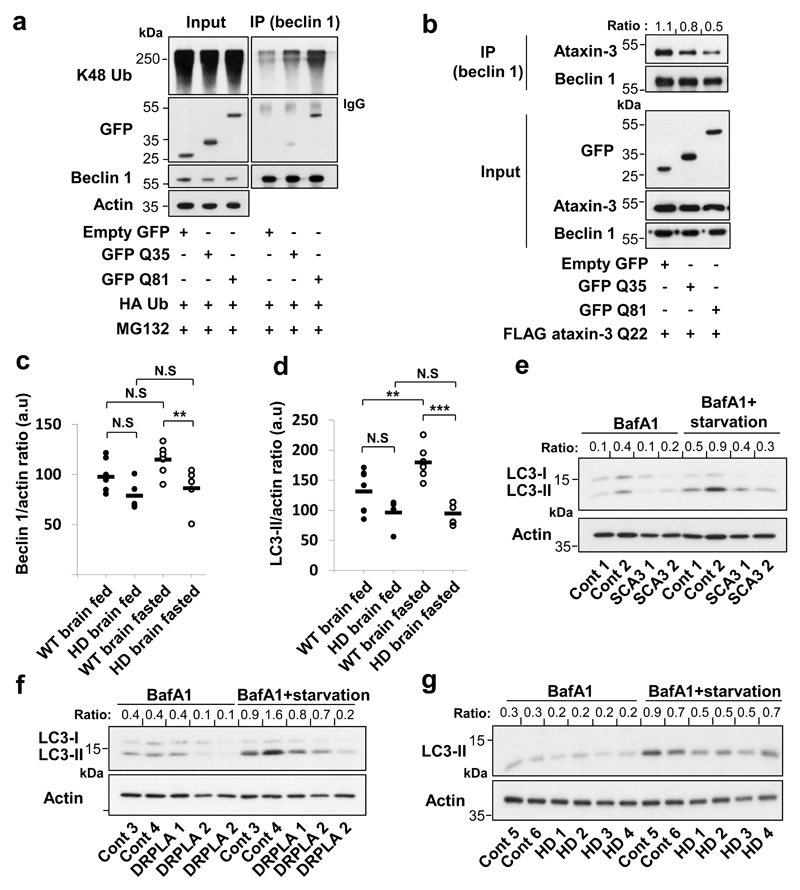
Nine neurodegenerative diseases are caused by expanded polyglutamine (polyQ) tracts in different proteins, such as huntingtin in Huntington's disease and ataxin 3 in spinocerebellar ataxia type 3 (SCA3). Age at onset of disease decreases with increasing polyglutamine length in these proteins and the normal length also varies. PolyQ expansions drive pathogenesis in these diseases, as isolated polyQ tracts are toxic, and an N-terminal huntingtin fragment comprising exon 1, which occurs in vivo as a result of alternative splicing, causes toxicity. Although such mutant proteins are prone to aggregation, toxicity is also associated with soluble forms of the proteins. The function of the polyQ tracts in many normal cytoplasmic proteins is unclear. One such protein is the deubiquitinating enzyme ataxin 3 (refs 7, 8), which is widely expressed in the brain. Here we show that the polyQ domain enables wild-type ataxin 3 to interact with beclin 1, a key initiator of autophagy. This interaction allows the deubiquitinase activity of ataxin 3 to protect beclin 1 from proteasome-mediated degradation and thereby enables autophagy. Starvation-induced autophagy, which is regulated by beclin 1, was particularly inhibited in ataxin-3-depleted human cell lines and mouse primary neurons, and in vivo in mice. This activity of ataxin 3 and its polyQ-mediated interaction with beclin 1 was competed for by other soluble proteins with polyQ tracts in a length-dependent fashion. This competition resulted in impairment of starvation-induced autophagy in cells expressing mutant huntingtin exon 1, and this impairment was recapitulated in the brains of a mouse model of Huntington's disease and in cells from patients. A similar phenomenon was also seen with other polyQ disease proteins, including mutant ataxin 3 itself. Our data thus describe a specific function for a wild-type polyQ tract that is abrogated by a competing longer polyQ mutation in a disease protein, and identify a deleterious function of such mutations distinct from their propensity to aggregate.
Conflict of interest statement
Potential competing financial interests: F.M.M. is currently an employee of Eli Lilly & Co. Ltd.
Figures
Comment in
-
Neurodegeneration: Role of repeats in protein clearance.Nature. 2017 May 4;545(7652):33-34. doi: 10.1038/nature22489. Epub 2017 Apr 26. Nature. 2017. PMID: 28445458 No abstract available.
-
Mechanisms of diseases: Excessive polyQ tracts curb autophagy.Nat Rev Mol Cell Biol. 2017 Jun;18(6):344. doi: 10.1038/nrm.2017.50. Epub 2017 May 10. Nat Rev Mol Cell Biol. 2017. PMID: 28488702 No abstract available.
-
Long polyglutamine tracts impair the autophagy protein quality control pathway.Mov Disord. 2017 Oct;32(10):1376. doi: 10.1002/mds.27118. Epub 2017 Aug 7. Mov Disord. 2017. PMID: 28782847 No abstract available.
Similar articles
-
Polyglutamine tracts regulate autophagy.Autophagy. 2017 Sep 2;13(9):1613-1614. doi: 10.1080/15548627.2017.1336278. Epub 2017 Jul 19. Autophagy. 2017. PMID: 28722507 Free PMC article.
-
Toward therapeutic targets for SCA3: Insight into the role of Machado-Joseph disease protein ataxin-3 in misfolded proteins clearance.Prog Neurobiol. 2015 Sep;132:34-58. doi: 10.1016/j.pneurobio.2015.06.004. Epub 2015 Jun 27. Prog Neurobiol. 2015. PMID: 26123252 Review.
-
Praja1 ubiquitin ligase facilitates degradation of polyglutamine proteins and suppresses polyglutamine-mediated toxicity.Mol Biol Cell. 2021 Aug 15;32(17):1579-1593. doi: 10.1091/mbc.E20-11-0747. Epub 2021 Jun 23. Mol Biol Cell. 2021. PMID: 34161122 Free PMC article.
-
Polyglutamine expansion diseases: More than simple repeats.J Struct Biol. 2018 Feb;201(2):139-154. doi: 10.1016/j.jsb.2017.09.006. Epub 2017 Sep 18. J Struct Biol. 2018. PMID: 28928079 Review.
-
Allele-selective lowering of mutant HTT protein by HTT-LC3 linker compounds.Nature. 2019 Nov;575(7781):203-209. doi: 10.1038/s41586-019-1722-1. Epub 2019 Oct 30. Nature. 2019. PMID: 31666698
Cited by
-
Small Molecules Inducing Autophagic Degradation of Expanded Polyglutamine Protein through Interaction with Both Mutant ATXN3 and LC3.Int J Mol Sci. 2024 Oct 4;25(19):10707. doi: 10.3390/ijms251910707. Int J Mol Sci. 2024. PMID: 39409036 Free PMC article.
-
Type IIa RPTPs and Glycans: Roles in Axon Regeneration and Synaptogenesis.Int J Mol Sci. 2021 May 24;22(11):5524. doi: 10.3390/ijms22115524. Int J Mol Sci. 2021. PMID: 34073798 Free PMC article. Review.
-
Age- and disease-related autophagy impairment in Huntington disease: New insights from direct neuronal reprogramming.Aging Cell. 2024 Aug;23(8):e14285. doi: 10.1111/acel.14285. Epub 2024 Jul 23. Aging Cell. 2024. PMID: 39044402 Free PMC article. Review.
-
Targeting AR-Beclin 1 complex-modulated growth factor signaling increases the antiandrogen Enzalutamide sensitivity to better suppress the castration-resistant prostate cancer growth.Cancer Lett. 2019 Feb 1;442:483-490. doi: 10.1016/j.canlet.2018.11.008. Epub 2018 Nov 10. Cancer Lett. 2019. PMID: 30423407 Free PMC article.
-
Leucine Signals to mTORC1 via Its Metabolite Acetyl-Coenzyme A.Cell Metab. 2019 Jan 8;29(1):192-201.e7. doi: 10.1016/j.cmet.2018.08.013. Epub 2018 Sep 6. Cell Metab. 2019. PMID: 30197302 Free PMC article.
References
-
- DiFiglia M, et al. Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science. 1997;277:1990–1993. - PubMed
-
- Riess O, Rub U, Pastore A, Bauer P, Schols L. SCA3: neurological features, pathogenesis and animal models. Cerebellum. 2008;7:125–137. - PubMed
-
- Imarisio S, et al. Huntington's disease: from pathology and genetics to potential therapies. The Biochemical journal. 2008;412:191–209. - PubMed
-
- Rubinsztein DC. The roles of intracellular protein-degradation pathways in neurodegeneration. Nature. 2006;443:780–786. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
