

Polycomb Repressive Complex 2-Mediated Chromatin Repression Guides Effector CD8+ T Cell Terminal Differentiation and Loss of Multipotency
- PMID: 28410989
- PMCID: PMC5457165
- DOI: 10.1016/j.immuni.2017.03.012
Polycomb Repressive Complex 2-Mediated Chromatin Repression Guides Effector CD8+ T Cell Terminal Differentiation and Loss of Multipotency
Abstract
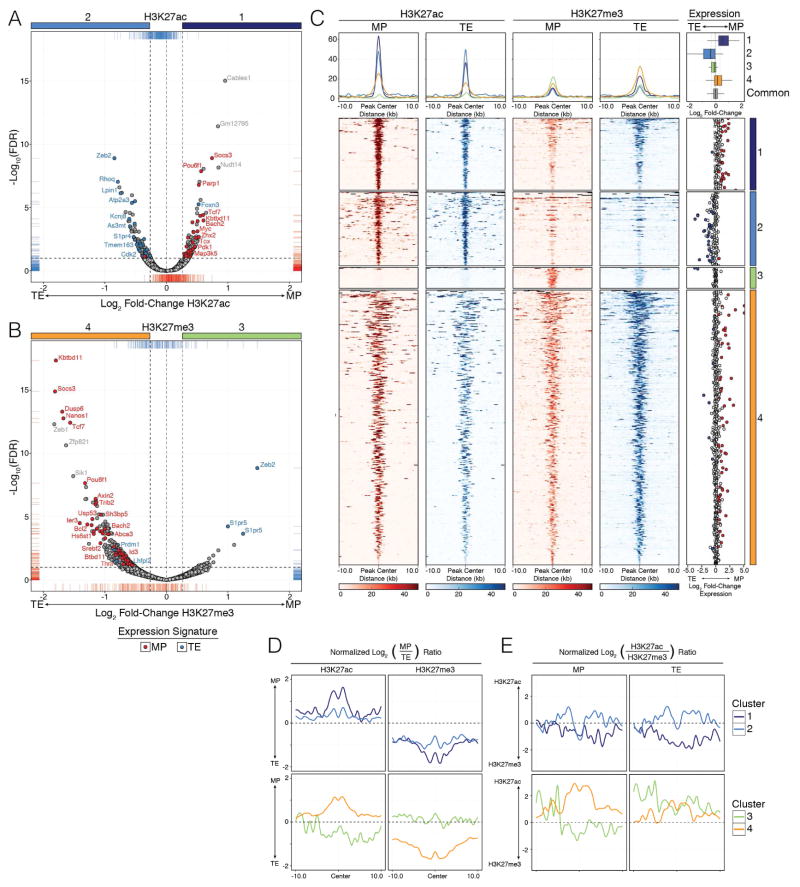
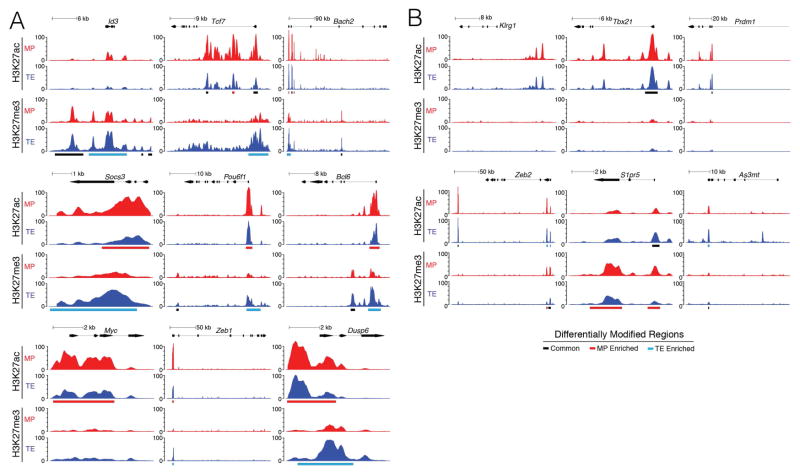
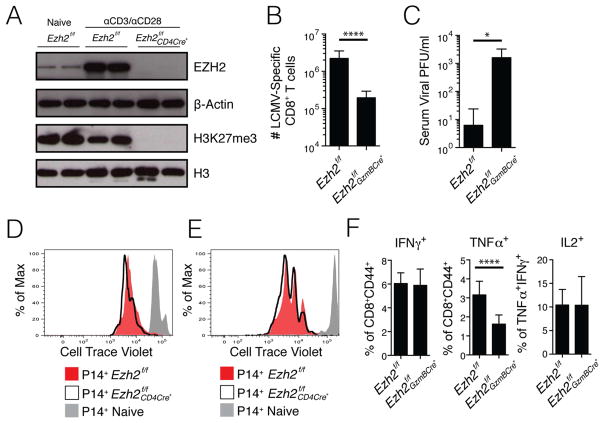
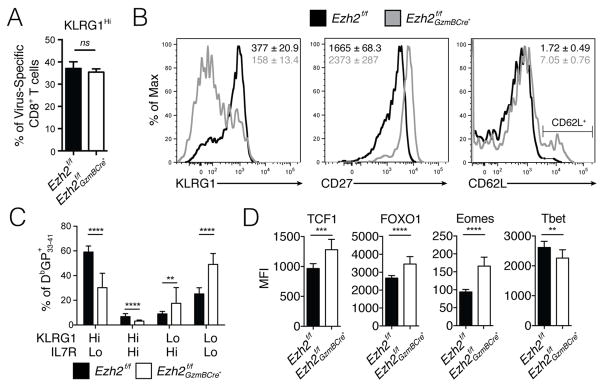
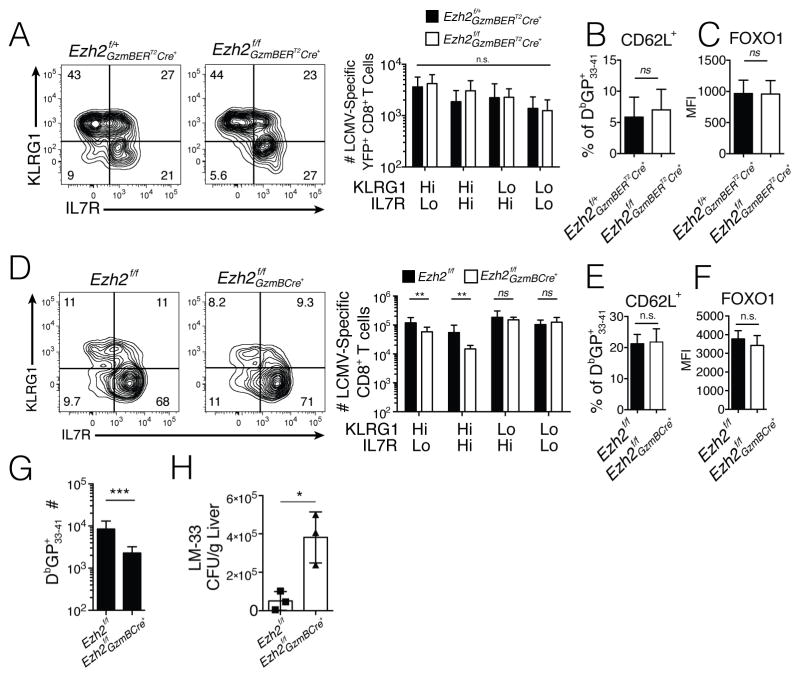
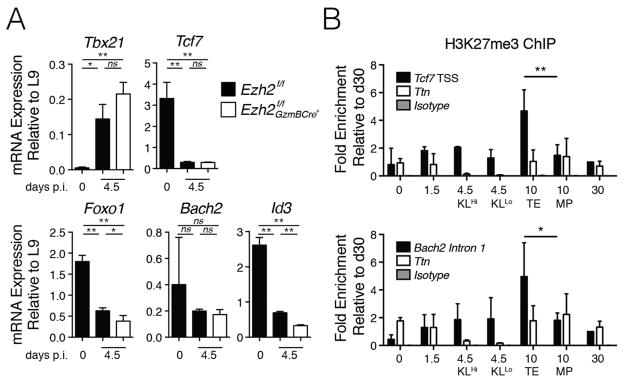
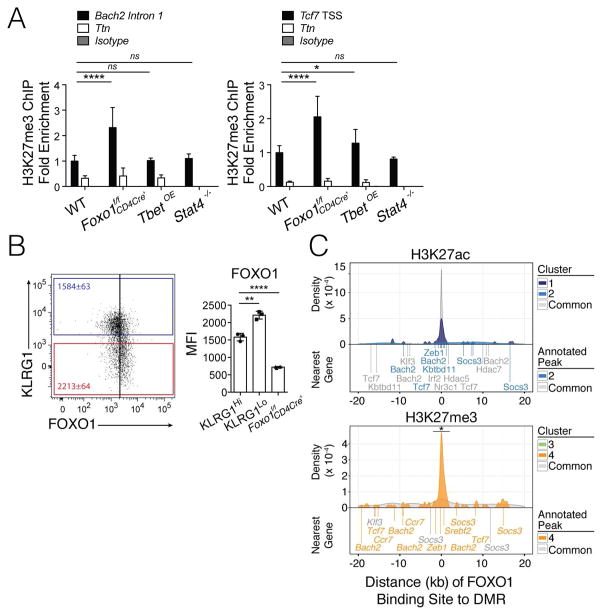
Understanding immunological memory formation depends on elucidating how multipotent memory precursor (MP) cells maintain developmental plasticity and longevity to provide long-term immunity while other effector cells develop into terminally differentiated effector (TE) cells with limited survival. Profiling active (H3K27ac) and repressed (H3K27me3) chromatin in naive, MP, and TE CD8+ T cells during viral infection revealed increased H3K27me3 deposition at numerous pro-memory and pro-survival genes in TE relative to MP cells, indicative of fate restriction, but permissive chromatin at both pro-memory and pro-effector genes in MP cells, indicative of multipotency. Polycomb repressive complex 2 deficiency impaired clonal expansion and TE cell differentiation, but minimally impacted CD8+ memory T cell maturation. Abundant H3K27me3 deposition at pro-memory genes occurred late during TE cell development, probably from diminished transcription factor FOXO1 expression. These results outline a temporal model for loss of memory cell potential through selective epigenetic silencing of pro-memory genes in effector T cells.
Keywords: CD8(+) T cell differentiation; EZH2; FOXO1; H3K27ac; H3K27me3; PRC2; Polycomb repressive complex 2; epigenetics; plasticity; terminal differentiation.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Polycomb Repressive Complex 2 Proteins EZH1 and EZH2 Regulate Timing of Postnatal Hepatocyte Maturation and Fibrosis by Repressing Genes With Euchromatic Promoters in Mice.Gastroenterology. 2019 May;156(6):1834-1848. doi: 10.1053/j.gastro.2019.01.041. Epub 2019 Jan 25. Gastroenterology. 2019. PMID: 30689973 Free PMC article.
-
Cooperation between the H3K27me3 Chromatin Mark and Non-CG Methylation in Epigenetic Regulation.Plant Physiol. 2016 Oct;172(2):1131-1141. doi: 10.1104/pp.16.01238. Epub 2016 Aug 17. Plant Physiol. 2016. PMID: 27535791 Free PMC article.
-
Ezh2 phosphorylation state determines its capacity to maintain CD8+ T memory precursors for antitumor immunity.Nat Commun. 2017 Dec 14;8(1):2125. doi: 10.1038/s41467-017-02187-8. Nat Commun. 2017. PMID: 29242551 Free PMC article.
-
Not just a writer: PRC2 as a chromatin reader.Biochem Soc Trans. 2021 Jun 30;49(3):1159-1170. doi: 10.1042/BST20200728. Biochem Soc Trans. 2021. PMID: 34060617 Free PMC article. Review.
-
EZH2 as a Regulator of CD8+ T Cell Fate and Function.Front Immunol. 2020 Oct 6;11:593203. doi: 10.3389/fimmu.2020.593203. eCollection 2020. Front Immunol. 2020. PMID: 33117406 Free PMC article. Review.
Cited by
-
A Single-Cell Perspective on Memory T-Cell Differentiation.Cold Spring Harb Perspect Biol. 2021 Sep 1;13(9):a038067. doi: 10.1101/cshperspect.a038067. Cold Spring Harb Perspect Biol. 2021. PMID: 33903160 Free PMC article. Review.
-
Knowledge-primed neural networks enable biologically interpretable deep learning on single-cell sequencing data.Genome Biol. 2020 Aug 3;21(1):190. doi: 10.1186/s13059-020-02100-5. Genome Biol. 2020. PMID: 32746932 Free PMC article.
-
Single-cell analysis by mass cytometry reveals metabolic states of early-activated CD8+ T cells during the primary immune response.Immunity. 2021 Apr 13;54(4):829-844.e5. doi: 10.1016/j.immuni.2021.02.018. Epub 2021 Mar 10. Immunity. 2021. PMID: 33705706 Free PMC article.
-
Stem, Effector, and Hybrid States of Memory CD8+ T Cells.Trends Immunol. 2020 Jan;41(1):17-28. doi: 10.1016/j.it.2019.11.004. Epub 2019 Dec 4. Trends Immunol. 2020. PMID: 31810790 Free PMC article. Review.
-
Remembering to remember: T cell memory maintenance and plasticity.Curr Opin Immunol. 2019 Jun;58:89-97. doi: 10.1016/j.coi.2019.04.009. Epub 2019 Jun 3. Curr Opin Immunol. 2019. PMID: 31170601 Free PMC article. Review.
References
-
- Araki Y, Wang Z, Zang C, Wood WH, 3rd, Schones D, Cui K, Roh TY, Lhotsky B, Wersto RP, Peng W, Becker KG, Zhao K, Weng NP. Genome-wide analysis of histone methylation reveals chromatin state-based regulation of gene transcription and function of memory CD8+ T cells. Immunity. 2009;30:912–25. - PMC - PubMed
-
- Chang JT, Ciocca ML, Kinjyo I, Palanivel VR, Mcclurkin CE, Dejong CS, Mooney EC, Kim JS, Steinel NC, Oliaro J, Yin CC, Florea BI, Overkleeft HS, Berg LJ, Russell SM, Koretzky GA, Jordan MS, Reiner SL. Asymmetric proteasome segregation as a mechanism for unequal partitioning of the transcription factor T-bet during T lymphocyte division. Immunity. 2011;34:492–504. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
