

A microRNA family exerts maternal control on sex determination in C. elegans
- PMID: 28279983
- PMCID: PMC5358761
- DOI: 10.1101/gad.290155.116
A microRNA family exerts maternal control on sex determination in C. elegans
Abstract
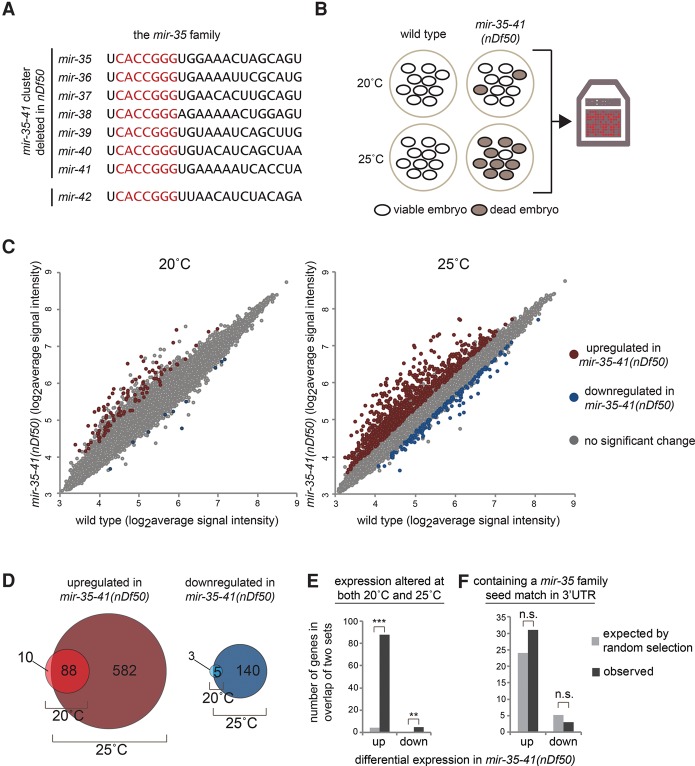
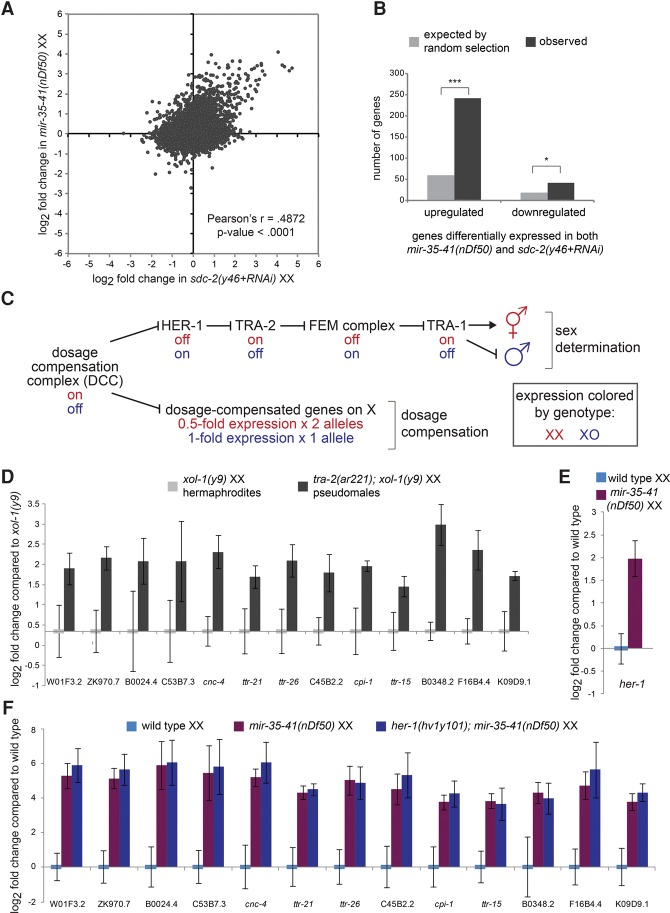
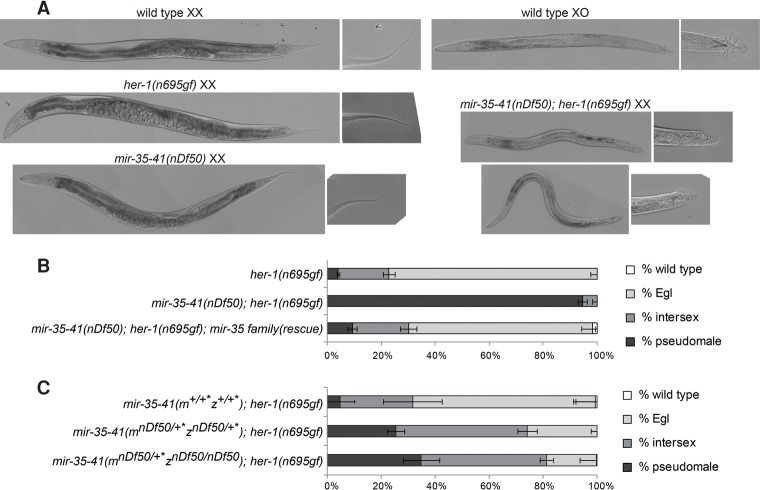
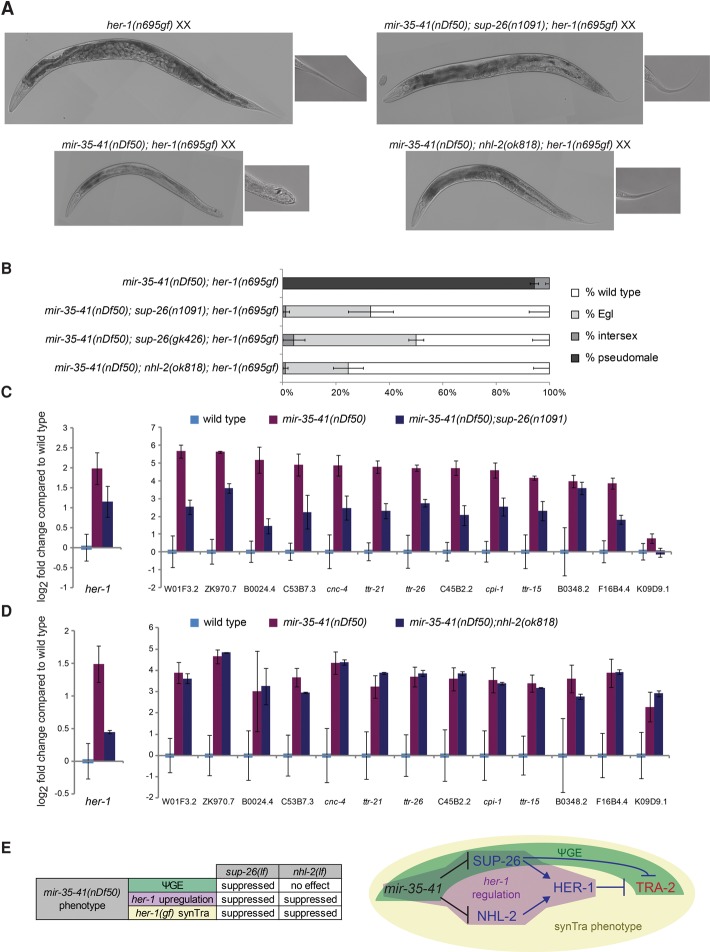
Gene expression in early animal embryogenesis is in large part controlled post-transcriptionally. Maternally contributed microRNAs may therefore play important roles in early development. We elucidated a major biological role of the nematode mir-35 family of maternally contributed essential microRNAs. We show that this microRNA family regulates the sex determination pathway at multiple levels, acting both upstream of and downstream from her-1 to prevent aberrantly activated male developmental programs in hermaphrodite embryos. Both of the predicted target genes that act downstream from the mir-35 family in this process, suppressor-26 (sup-26) and NHL (NCL-1, HT2A, and LIN-41 repeat) domain-containing-2 (nhl-2), encode RNA-binding proteins, thus delineating a previously unknown post-transcriptional regulatory subnetwork within the well-studied sex determination pathway of Caenorhabditis elegans Repression of nhl-2 by the mir-35 family is required for not only proper sex determination but also viability, showing that a single microRNA target site can be essential. Since sex determination in C. elegans requires zygotic gene expression to read the sex chromosome karyotype, early embryos must remain gender-naïve; our findings show that the mir-35 family microRNAs act in the early embryo to function as a developmental timer that preserves naïveté and prevents premature deleterious developmental decisions.
Keywords: embryonic development; maternal control; microRNAs; mir-35–41; mir-35–42; sex determination.
© 2017 McJunkin and Ambros; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Maternal effects of microRNAs in early embryogenesis.RNA Biol. 2018 Feb 1;15(2):165-169. doi: 10.1080/15476286.2017.1402999. Epub 2017 Dec 8. RNA Biol. 2018. PMID: 29120257 Free PMC article. Review.
-
The mir-35 Family Links Maternal Germline Sex to Embryonic Viability in Caenorhabditis elegans.G3 (Bethesda). 2019 Mar 7;9(3):901-909. doi: 10.1534/g3.118.200863. G3 (Bethesda). 2019. PMID: 30679246 Free PMC article.
-
Stage-Specific Timing of the microRNA Regulation of lin-28 by the Heterochronic Gene lin-14 in Caenorhabditis elegans.Genetics. 2017 Jan;205(1):251-262. doi: 10.1534/genetics.116.195040. Epub 2016 Nov 4. Genetics. 2017. PMID: 27815363 Free PMC article.
-
GW182-Free microRNA Silencing Complex Controls Post-transcriptional Gene Expression during Caenorhabditis elegans Embryogenesis.PLoS Genet. 2016 Dec 9;12(12):e1006484. doi: 10.1371/journal.pgen.1006484. eCollection 2016 Dec. PLoS Genet. 2016. PMID: 27935964 Free PMC article.
-
Roles of microRNAs in the Caenorhabditis elegans nervous system.J Genet Genomics. 2013 Sep 20;40(9):445-52. doi: 10.1016/j.jgg.2013.07.002. Epub 2013 Aug 7. J Genet Genomics. 2013. PMID: 24053946 Review.
Cited by
-
The mir-35-42 binding site in the nhl-2 3'UTR is dispensable for development and fecundity.MicroPubl Biol. 2020 Apr 14;2020:10.17912/micropub.biology.000241. doi: 10.17912/micropub.biology.000241. MicroPubl Biol. 2020. PMID: 32550481 Free PMC article. No abstract available.
-
A Compilation of the Diverse miRNA Functions in Caenorhabditis elegans and Drosophila melanogaster Development.Int J Mol Sci. 2023 Apr 9;24(8):6963. doi: 10.3390/ijms24086963. Int J Mol Sci. 2023. PMID: 37108126 Free PMC article. Review.
-
LIN-41 and OMA Ribonucleoprotein Complexes Mediate a Translational Repression-to-Activation Switch Controlling Oocyte Meiotic Maturation and the Oocyte-to-Embryo Transition in Caenorhabditis elegans.Genetics. 2017 Aug;206(4):2007-2039. doi: 10.1534/genetics.117.203174. Epub 2017 Jun 1. Genetics. 2017. PMID: 28576864 Free PMC article.
-
In vivo CRISPR screening for phenotypic targets of the mir-35-42 family in C. elegans.Genes Dev. 2020 Sep 1;34(17-18):1227-1238. doi: 10.1101/gad.339333.120. Epub 2020 Aug 20. Genes Dev. 2020. PMID: 32820039 Free PMC article.
-
Identification and Characterization of MicroRNAs Involving in Initial Sex Differentiation of Chlamys farreri Gonads.Biology (Basel). 2022 Mar 16;11(3):456. doi: 10.3390/biology11030456. Biology (Basel). 2022. PMID: 35336829 Free PMC article.
References
-
- Chuck G, Meeley R, Irish E, Sakai H, Hake S. 2007. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat Genet 39: 1517–1521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous