

The Chemokine Receptor CX3CR1 Defines Three Antigen-Experienced CD8 T Cell Subsets with Distinct Roles in Immune Surveillance and Homeostasis
- PMID: 27939671
- PMCID: PMC5177508
- DOI: 10.1016/j.immuni.2016.10.018
The Chemokine Receptor CX3CR1 Defines Three Antigen-Experienced CD8 T Cell Subsets with Distinct Roles in Immune Surveillance and Homeostasis
Abstract
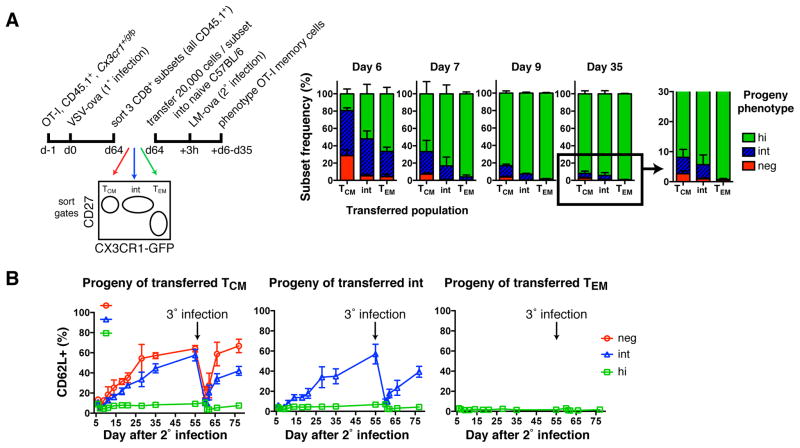
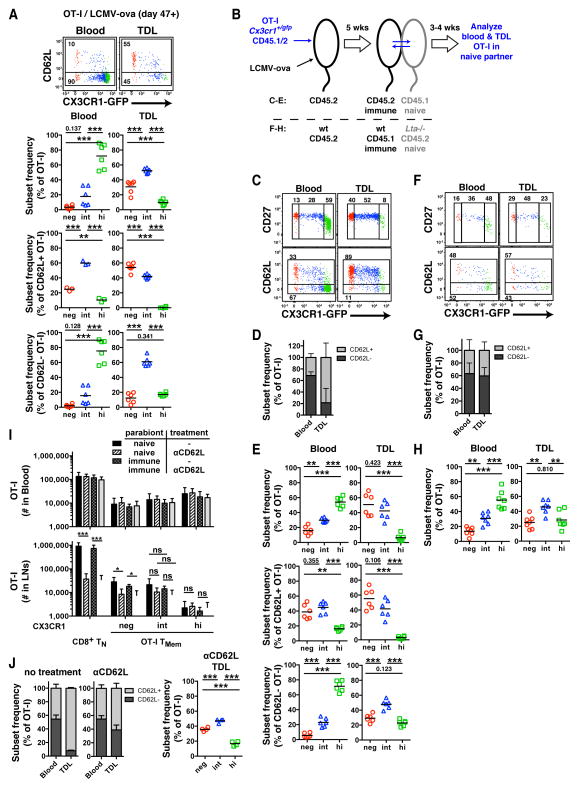
Infections induce pathogen-specific T cell differentiation into diverse effectors (Teff) that give rise to memory (Tmem) subsets. The cell-fate decisions and lineage relationships that underlie these transitions are poorly understood. Here, we found that the chemokine receptor CX3CR1 identifies three distinct CD8+ Teff and Tmem subsets. Classical central (Tcm) and effector memory (Tem) cells and their corresponding Teff precursors were CX3CR1- and CX3CR1high, respectively. Viral infection also induced a numerically stable CX3CR1int subset that represented ∼15% of blood-borne Tmem cells. CX3CR1int Tmem cells underwent more frequent homeostatic divisions than other Tmem subsets and not only self-renewed, but also contributed to the expanding CX3CR1- Tcm pool. Both Tcm and CX3CR1int cells homed to lymph nodes, but CX3CR1int cells, and not Tem cells, predominantly surveyed peripheral tissues. As CX3CR1int Tmem cells present unique phenotypic, homeostatic, and migratory properties, we designate this subset peripheral memory (tpm) cells and propose that tpm cells are chiefly responsible for the global surveillance of non-lymphoid tissues.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures





Comment in
-
Sifting through CD8+ T Cell Memory.Immunity. 2016 Dec 20;45(6):1184-1186. doi: 10.1016/j.immuni.2016.12.005. Immunity. 2016. PMID: 28002725
-
T cells: Sorting memories.Nat Rev Immunol. 2016 Dec 23;17(1):2-3. doi: 10.1038/nri.2016.146. Nat Rev Immunol. 2016. PMID: 28008185 No abstract available.
Similar articles
-
The chemokine receptor CX3CR1 controls homing and anti-viral potencies of CD8 effector-memory T lymphocytes in HIV-infected patients.AIDS. 2003 Jun 13;17(9):1279-90. doi: 10.1097/00002030-200306130-00002. AIDS. 2003. PMID: 12799549
-
Inflammatory monocytes contribute to the persistence of CXCR3hi CX3CR1lo circulating and lung-resident memory CD8+ T cells following respiratory virus infection.Immunol Cell Biol. 2018 Apr;96(4):370-378. doi: 10.1111/imcb.12006. Epub 2018 Jan 30. Immunol Cell Biol. 2018. PMID: 29363162 Free PMC article.
-
A Context-Dependent Role for IL-21 in Modulating the Differentiation, Distribution, and Abundance of Effector and Memory CD8 T Cell Subsets.J Immunol. 2016 Mar 1;196(5):2153-66. doi: 10.4049/jimmunol.1401236. Epub 2016 Jan 29. J Immunol. 2016. PMID: 26826252 Free PMC article.
-
Chemokine-mediated control of T cell traffic in lymphoid and peripheral tissues.Mol Immunol. 2005 May;42(7):799-809. doi: 10.1016/j.molimm.2004.06.040. Epub 2004 Nov 23. Mol Immunol. 2005. PMID: 15829268 Review.
-
Stem cell-like plasticity of naïve and distinct memory CD8+ T cell subsets.Semin Immunol. 2009 Apr;21(2):62-8. doi: 10.1016/j.smim.2009.02.004. Epub 2009 Mar 9. Semin Immunol. 2009. PMID: 19269852 Review.
Cited by
-
A mechanism for BCG vaccine cross-protection against pulmonary viral infections.Nat Immunol. 2024 Mar;25(3):403-404. doi: 10.1038/s41590-024-01775-3. Nat Immunol. 2024. PMID: 38383613 No abstract available.
-
Specific targeting of IL-1β activity to CD8+ T cells allows for safe use as a vaccine adjuvant.NPJ Vaccines. 2020 Jul 23;5(1):64. doi: 10.1038/s41541-020-00211-5. eCollection 2020. NPJ Vaccines. 2020. PMID: 32714571 Free PMC article.
-
Motility Matters: How CD8+ T-Cell Trafficking Influences Effector and Memory Cell Differentiation.Cold Spring Harb Perspect Biol. 2021 Aug 2;13(8):a038075. doi: 10.1101/cshperspect.a038075. Cold Spring Harb Perspect Biol. 2021. PMID: 34001529 Free PMC article. Review.
-
The Memory T Cell "Communication Web" in Context with Gastrointestinal Disorders-How Memory T Cells Affect Their Surroundings and How They Are Influenced by It.Cells. 2022 Sep 6;11(18):2780. doi: 10.3390/cells11182780. Cells. 2022. PMID: 36139354 Free PMC article. Review.
-
Long-lived central memory γδ T cells confer protection against murine cytomegalovirus reinfection.PLoS Pathog. 2024 Jul 8;20(7):e1010785. doi: 10.1371/journal.ppat.1010785. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 38976755 Free PMC article.
References
-
- Bargatze RF, Jutila MA, Butcher EC. Distinct roles of L-selectin and integrins α4β7 and LFA-1 in lymphocyte homing to Peyer’s patch-HEV in situ: The multistep model confirmed and refined. Immunity. 1995;3:99–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
