

How and why intralumenal membrane fragments form during vacuolar lysosome fusion
- PMID: 27881666
- PMCID: PMC5231899
- DOI: 10.1091/mbc.E15-11-0759
How and why intralumenal membrane fragments form during vacuolar lysosome fusion
Abstract
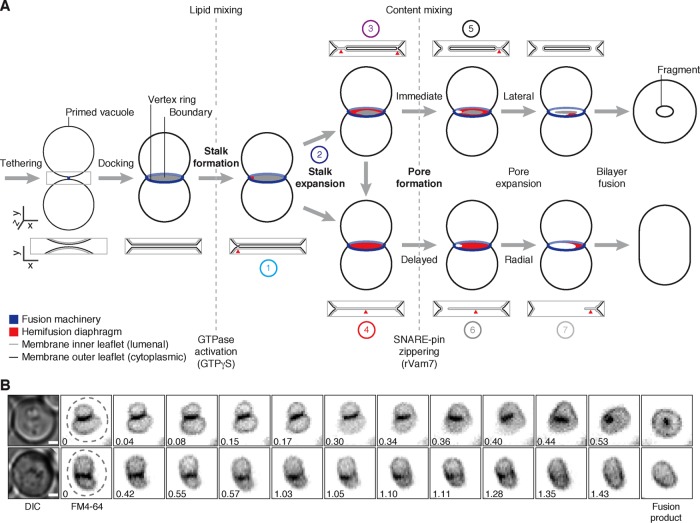
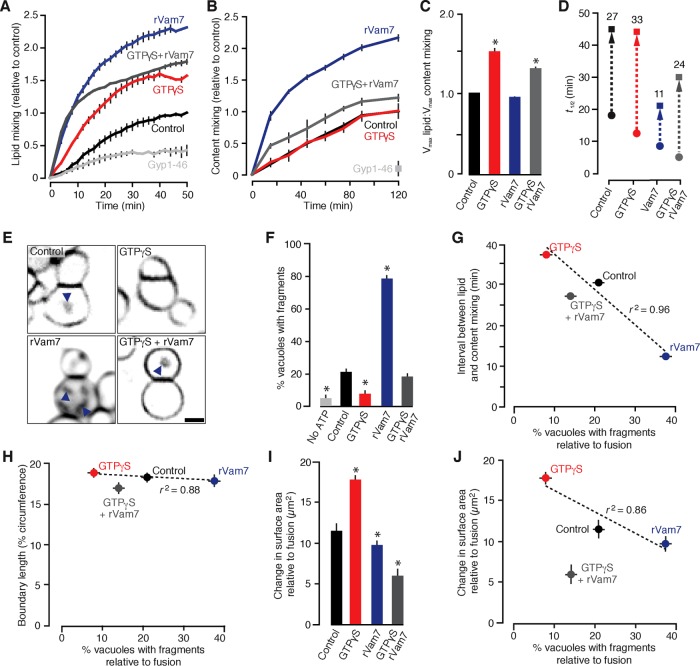
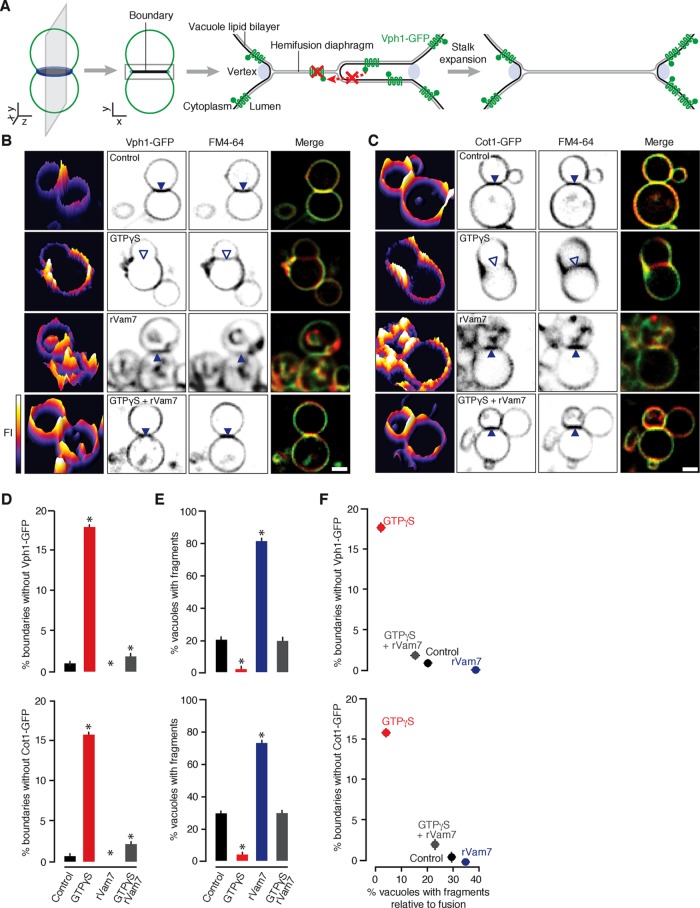
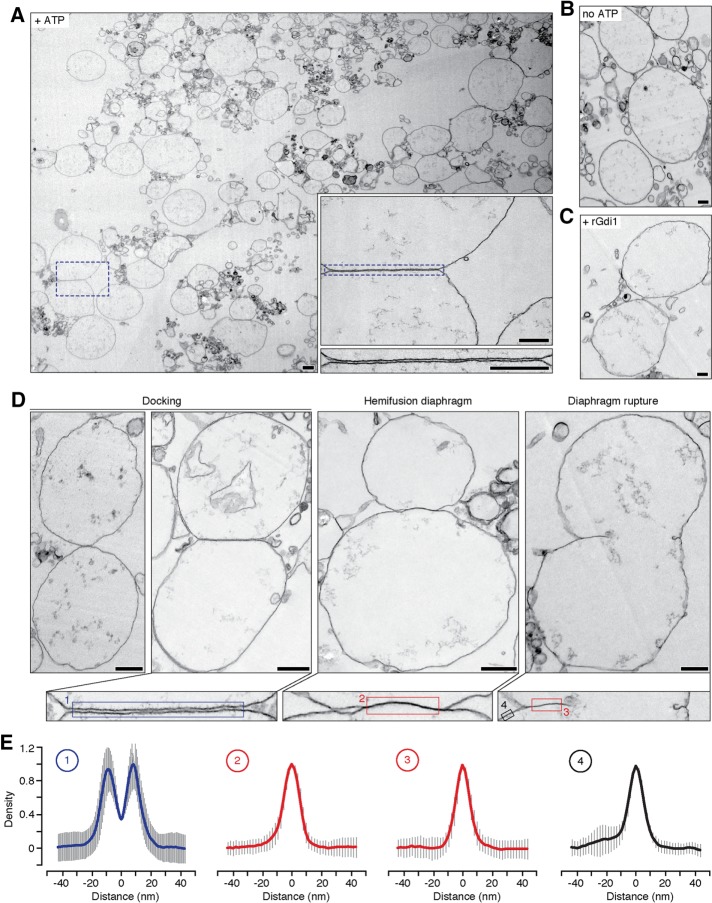
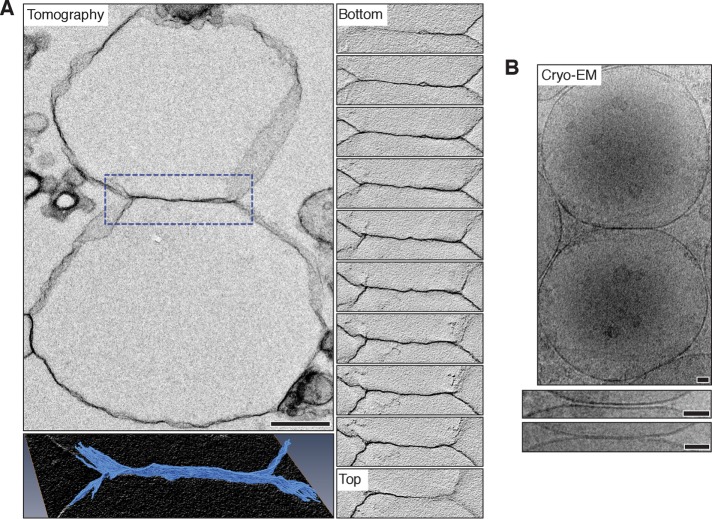
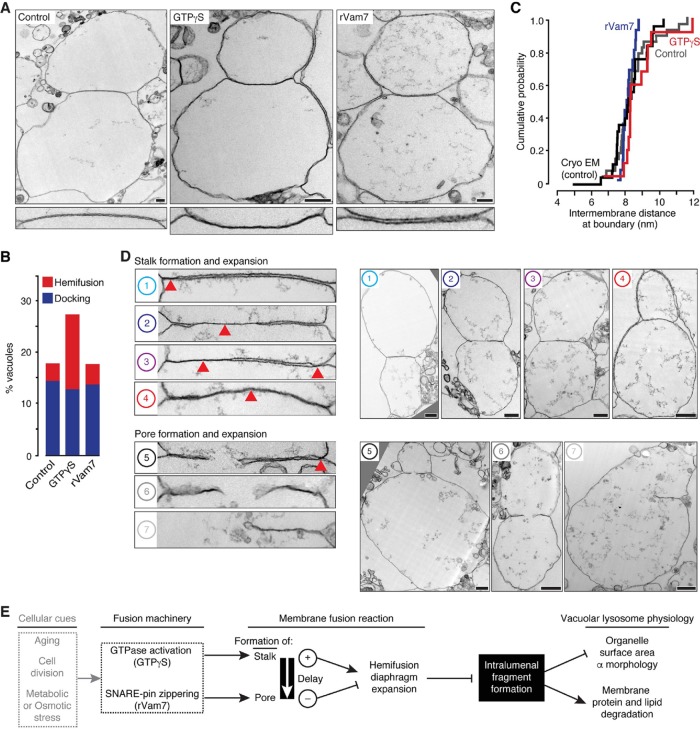
Lysosomal membrane fusion mediates the last step of the autophagy and endocytosis pathways and supports organelle remodeling and biogenesis. Because fusogenic proteins and lipids concentrate in a ring at the vertex between apposing organelle membranes, the encircled area of membrane can be severed and internalized within the lumen as a fragment upon lipid bilayer fusion. How or why this intralumenal fragment forms during fusion, however, is not entirely clear. To better understand this process, we studied fragment formation during homotypic vacuolar lysosome membrane fusion in Saccharomyces cerevisiae Using cell-free fusion assays and light microscopy, we find that GTPase activation and trans-SNARE complex zippering have opposing effects on fragment formation and verify that this affects the morphology of the fusion product and regulates transporter protein degradation. We show that fragment formwation is limited by stalk expansion, a key intermediate of the lipid bilayer fusion reaction. Using electron microscopy, we present images of hemifusion diaphragms that form as stalks expand and propose a model describing how the fusion machinery regulates fragment formation during lysosome fusion to control morphology and protein lifetimes.
© 2017 Mattie et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
Similar articles
-
Rab-Effector-Kinase Interplay Modulates Intralumenal Fragment Formation during Vacuole Fusion.Dev Cell. 2018 Oct 8;47(1):80-97.e6. doi: 10.1016/j.devcel.2018.09.002. Epub 2018 Sep 27. Dev Cell. 2018. PMID: 30269949
-
Visualization of SNARE-Mediated Organelle Membrane Hemifusion by Electron Microscopy.Methods Mol Biol. 2019;1860:361-377. doi: 10.1007/978-1-4939-8760-3_24. Methods Mol Biol. 2019. PMID: 30317518
-
Selective Lysosomal Transporter Degradation by Organelle Membrane Fusion.Dev Cell. 2017 Jan 23;40(2):151-167. doi: 10.1016/j.devcel.2016.11.024. Epub 2016 Dec 22. Dev Cell. 2017. PMID: 28017618
-
Yeast vacuole fusion: a model system for eukaryotic endomembrane dynamics.Autophagy. 2008 Jan;4(1):5-19. doi: 10.4161/auto.5054. Epub 2007 Sep 12. Autophagy. 2008. PMID: 17932463 Review.
-
Functions of SNAREs in intracellular membrane fusion and lipid bilayer mixing.J Cell Sci. 2005 Sep 1;118(Pt 17):3819-28. doi: 10.1242/jcs.02561. J Cell Sci. 2005. PMID: 16129880 Review.
Cited by
-
A hemifused complex is the hub in a network of pathways to membrane fusion.Biophys J. 2023 Jan 17;122(2):374-385. doi: 10.1016/j.bpj.2022.12.003. Epub 2022 Dec 5. Biophys J. 2023. PMID: 36463406 Free PMC article.
-
Membrane Fusion Mediated by Non-covalent Binding of Re-engineered Cholera Toxin Assemblies to Glycolipids.ACS Synth Biol. 2022 Dec 16;11(12):3929-3938. doi: 10.1021/acssynbio.2c00266. Epub 2022 Nov 11. ACS Synth Biol. 2022. PMID: 36367814 Free PMC article.
-
HOPS-dependent endosomal fusion required for efficient cytosolic delivery of therapeutic peptides and small proteins.Proc Natl Acad Sci U S A. 2019 Jan 8;116(2):512-521. doi: 10.1073/pnas.1812044116. Proc Natl Acad Sci U S A. 2019. PMID: 30610181 Free PMC article.
-
Evidence for ESCRT- and clathrin-dependent microautophagy.J Cell Biol. 2017 Oct 2;216(10):3263-3274. doi: 10.1083/jcb.201611029. Epub 2017 Aug 24. J Cell Biol. 2017. PMID: 28838958 Free PMC article.
-
SNARE-mediated membrane fusion arrests at pore expansion to regulate the volume of an organelle.EMBO J. 2018 Oct 1;37(19):e99193. doi: 10.15252/embj.201899193. Epub 2018 Aug 17. EMBO J. 2018. PMID: 30120144 Free PMC article.
References
-
- Berger AC, Hanson PK, Wylie Nichols J, Corbett AH. A yeast model system for functional analysis of the Niemann-Pick type C protein 1 homolog, Ncr1p. Traffic. 2005;6:907–917. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
