

Different tissue phagocytes sample apoptotic cells to direct distinct homeostasis programs
- PMID: 27828940
- PMCID: PMC5807003
- DOI: 10.1038/nature20138
Different tissue phagocytes sample apoptotic cells to direct distinct homeostasis programs
Abstract
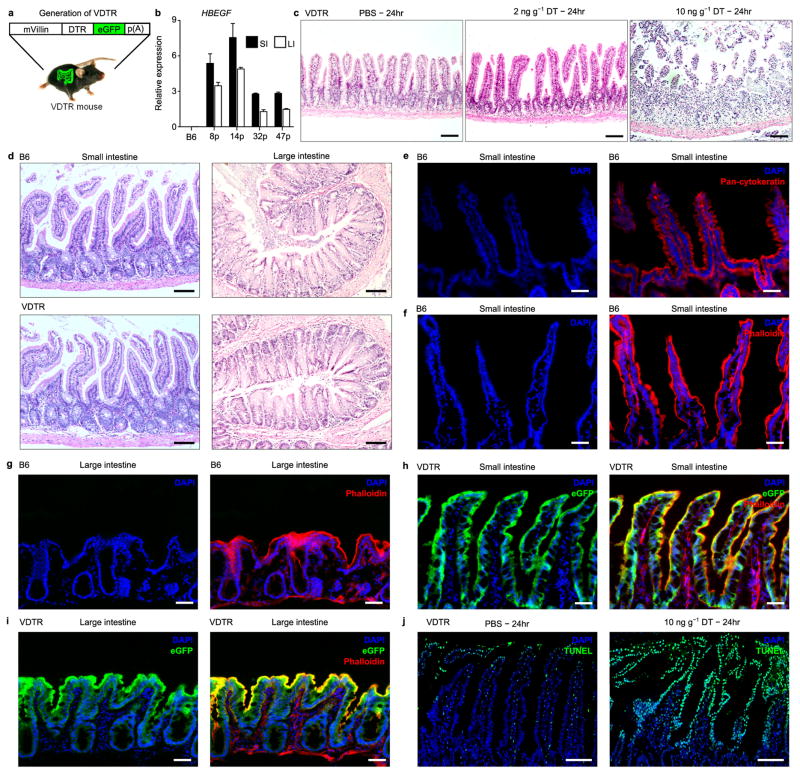
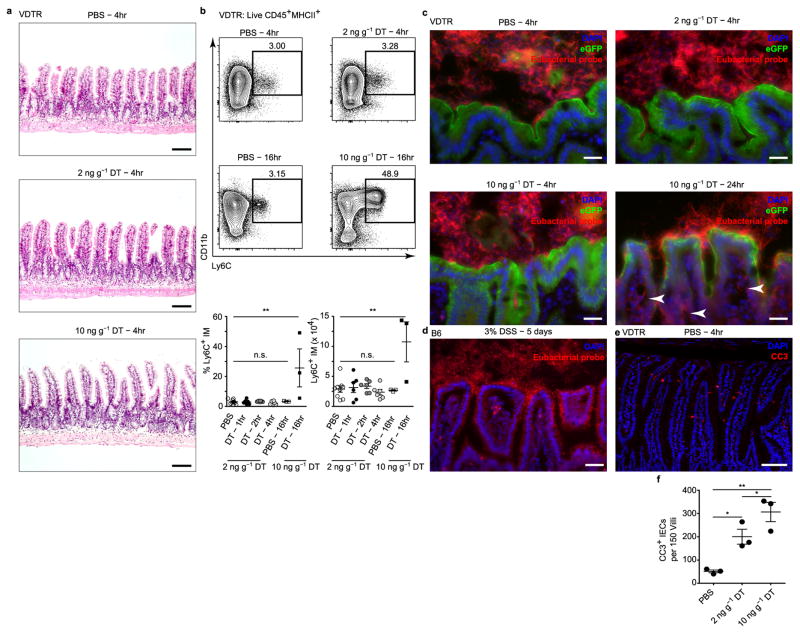
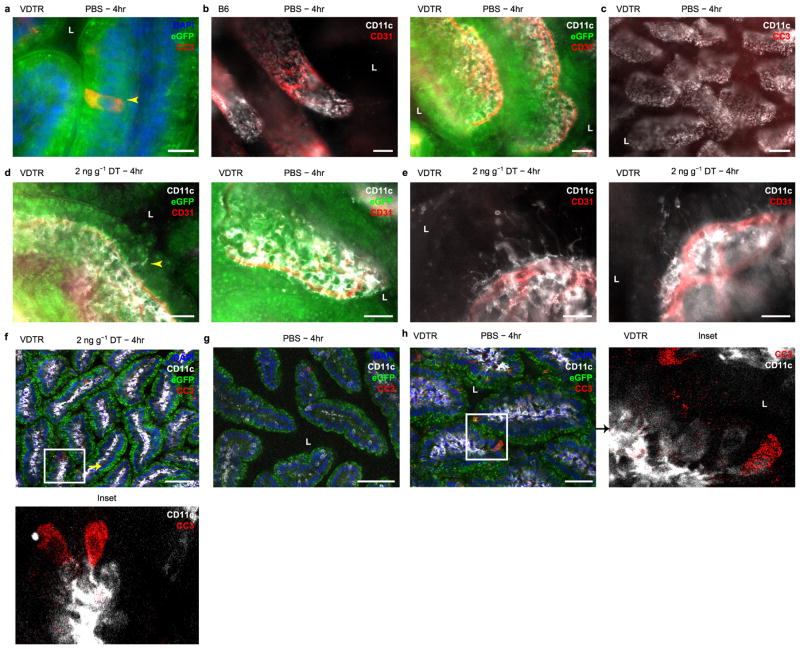
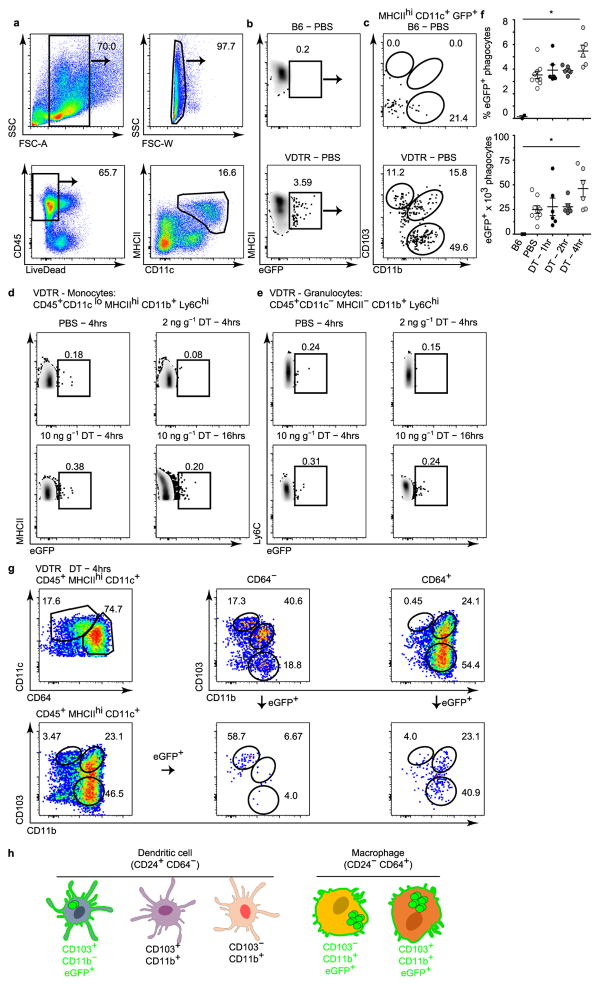
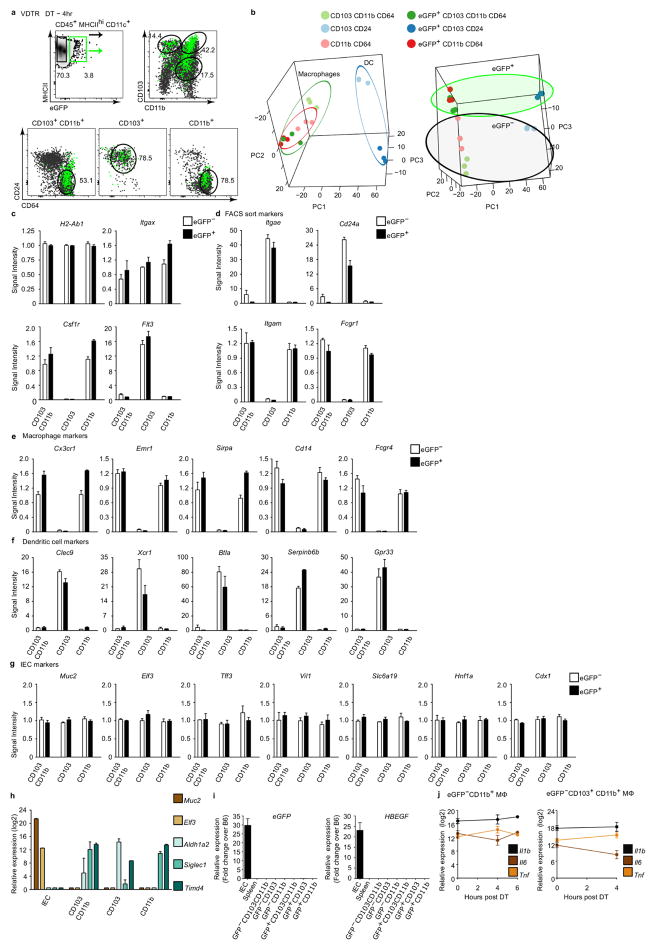
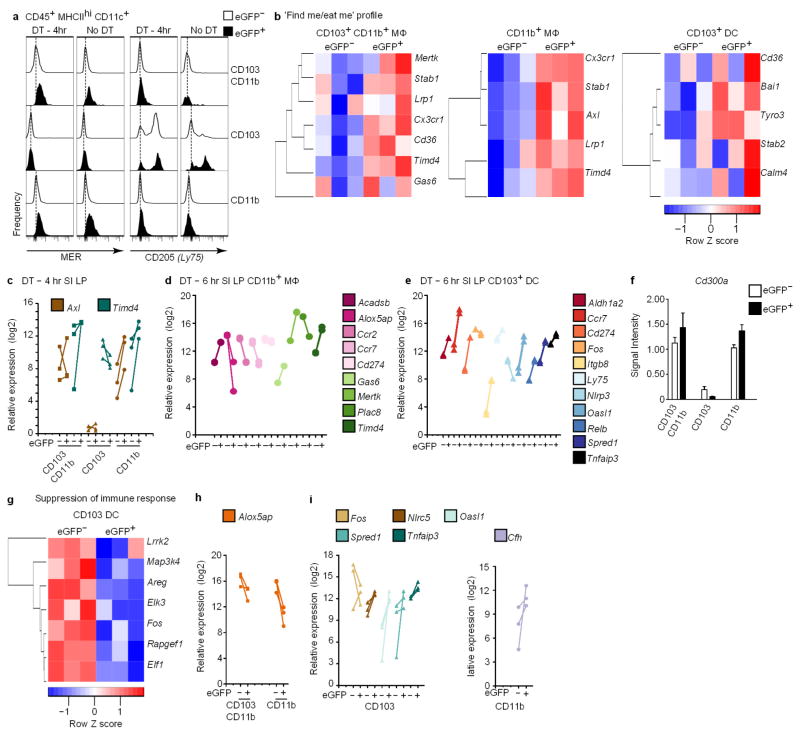
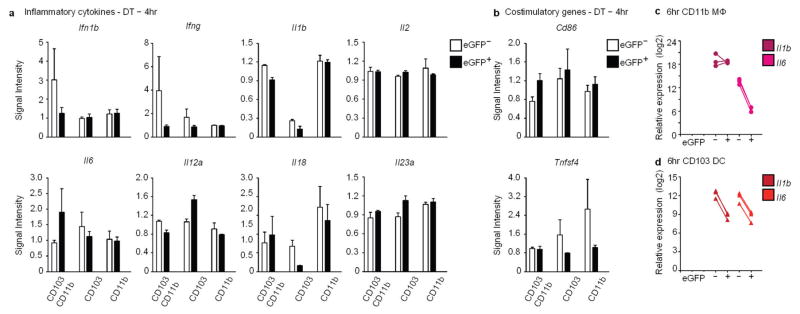
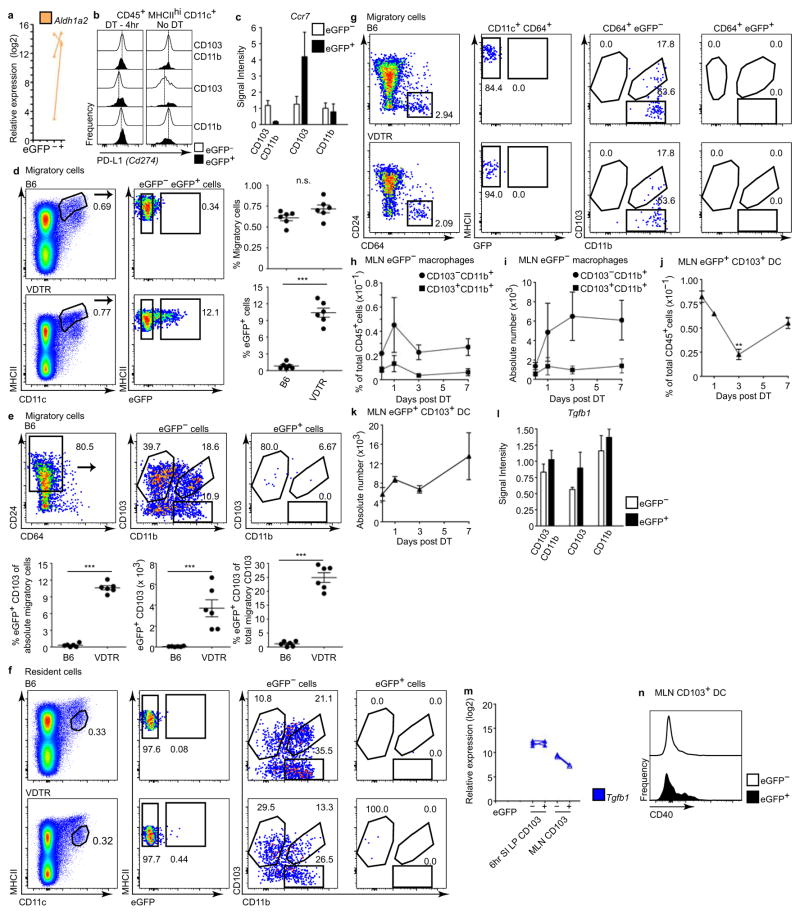
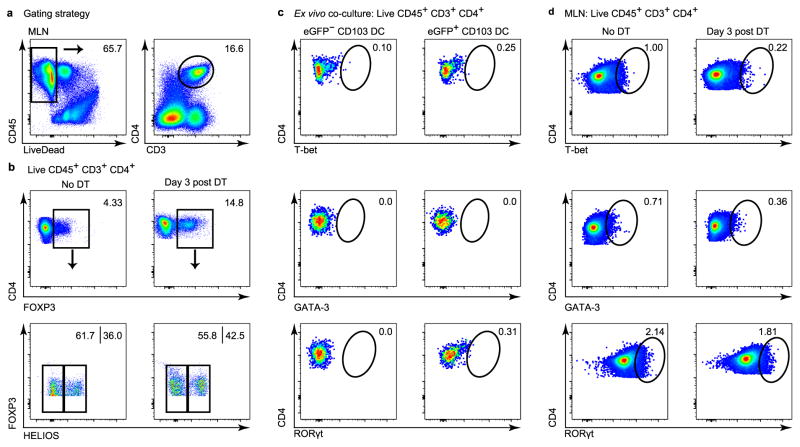
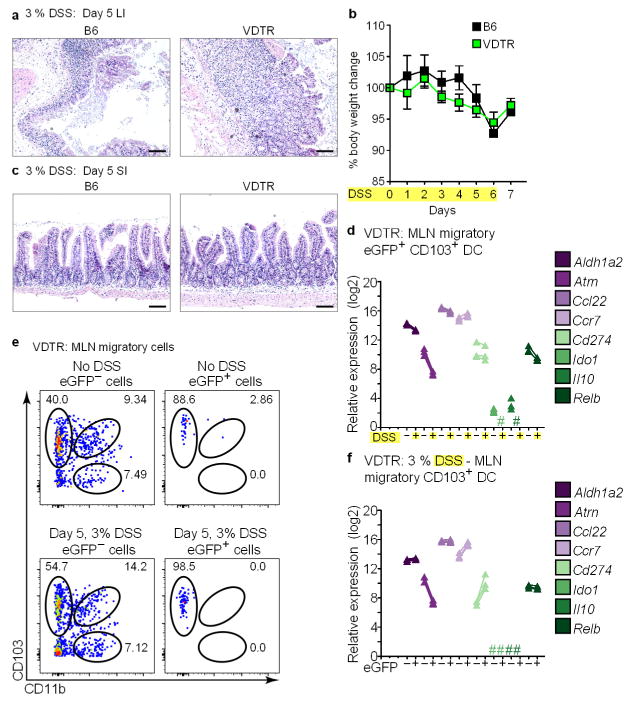
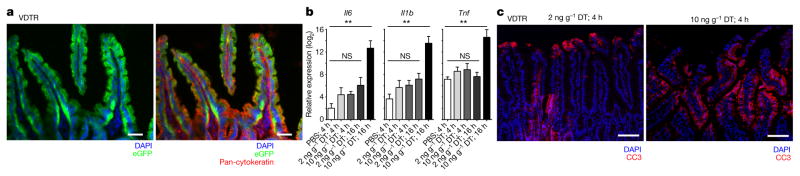
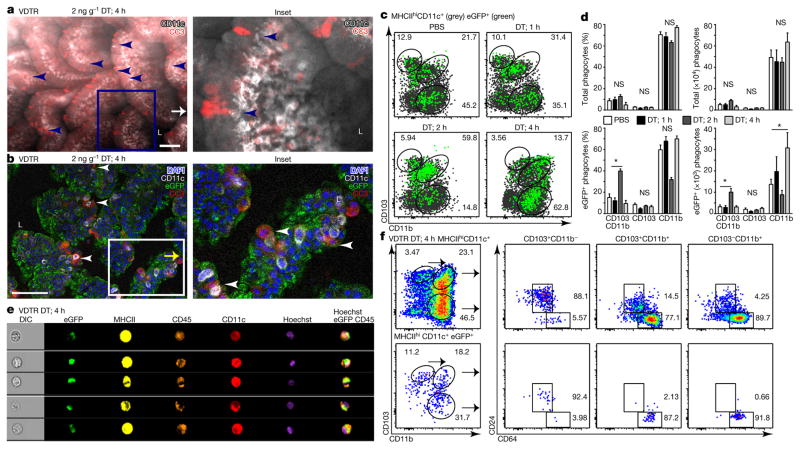
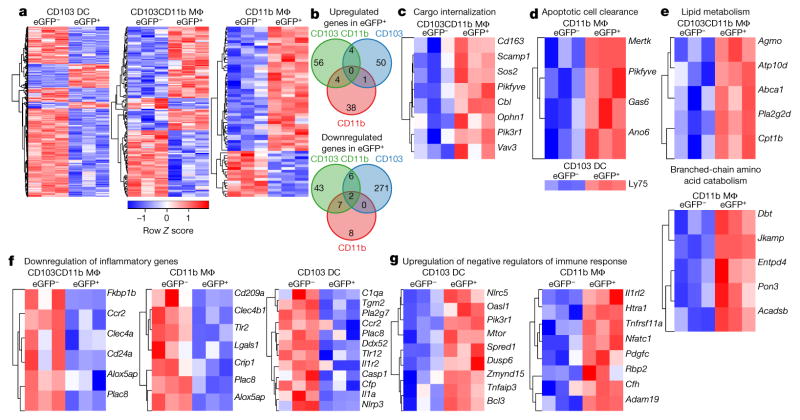
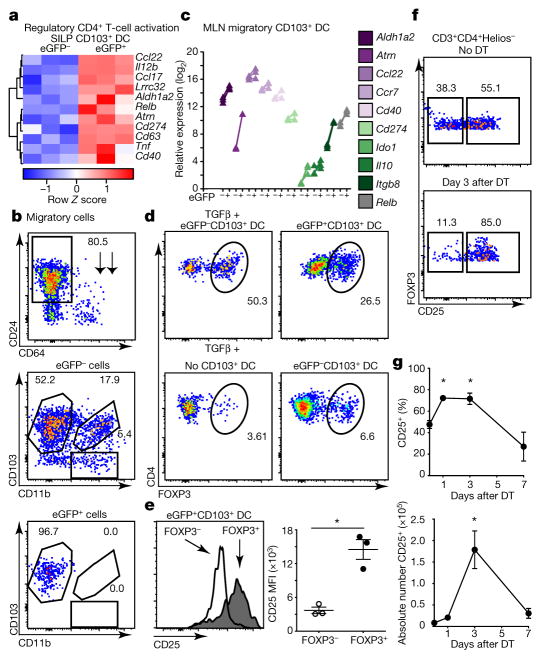
Recognition and removal of apoptotic cells by professional phagocytes, including dendritic cells and macrophages, preserves immune self-tolerance and prevents chronic inflammation and autoimmune pathologies. The diverse array of phagocytes that reside within different tissues, combined with the necessarily prompt nature of apoptotic cell clearance, makes it difficult to study this process in situ. The full spectrum of functions executed by tissue-resident phagocytes in response to homeostatic apoptosis, therefore, remains unclear. Here we show that mouse apoptotic intestinal epithelial cells (IECs), which undergo continuous renewal to maintain optimal barrier and absorptive functions, are not merely extruded to maintain homeostatic cell numbers, but are also sampled by a single subset of dendritic cells and two macrophage subsets within a well-characterized network of phagocytes in the small intestinal lamina propria. Characterization of the transcriptome within each subset before and after in situ sampling of apoptotic IECs revealed gene expression signatures unique to each phagocyte, including macrophage-specific lipid metabolism and amino acid catabolism, and a dendritic-cell-specific program of regulatory CD4+ T-cell activation. A common 'suppression of inflammation' signature was noted, although the specific genes and pathways involved varied amongst dendritic cells and macrophages, reflecting specialized functions. Apoptotic IECs were trafficked to mesenteric lymph nodes exclusively by the dendritic cell subset and served as critical determinants for the induction of tolerogenic regulatory CD4+ T-cell differentiation. Several of the genes that were differentially expressed by phagocytes bearing apoptotic IECs overlapped with susceptibility genes for inflammatory bowel disease. Collectively, these findings provide new insights into the consequences of apoptotic cell sampling, advance our understanding of how homeostasis is maintained within the mucosa and set the stage for development of novel therapeutics to alleviate chronic inflammatory diseases such as inflammatory bowel disease.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Gut check: dead cell samples leads to tolerant examples.Cell Death Differ. 2017 Sep;24(9):1471-1472. doi: 10.1038/cdd.2017.45. Epub 2017 Jun 23. Cell Death Differ. 2017. PMID: 28644438 Free PMC article. No abstract available.
Similar articles
-
On cell death in the intestinal epithelium and its impact on gut homeostasis.Curr Opin Gastroenterol. 2018 Nov;34(6):413-419. doi: 10.1097/MOG.0000000000000481. Curr Opin Gastroenterol. 2018. PMID: 30169459 Free PMC article. Review.
-
The many ways tissue phagocytes respond to dying cells.Immunol Rev. 2017 May;277(1):158-173. doi: 10.1111/imr.12537. Immunol Rev. 2017. PMID: 28462530 Free PMC article. Review.
-
CX3CR1⁺ cells facilitate the activation of CD4 T cells in the colonic lamina propria during antigen-driven colitis.Mucosal Immunol. 2014 May;7(3):533-48. doi: 10.1038/mi.2013.70. Epub 2013 Oct 16. Mucosal Immunol. 2014. PMID: 24129164
-
Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells.Gut. 2009 Nov;58(11):1481-9. doi: 10.1136/gut.2008.175166. Epub 2009 Jun 30. Gut. 2009. PMID: 19570762
-
Immune microniches shape intestinal Treg function.Nature. 2024 Apr;628(8009):854-862. doi: 10.1038/s41586-024-07251-0. Epub 2024 Apr 3. Nature. 2024. PMID: 38570678 Free PMC article.
Cited by
-
TNF-α sculpts a maturation process in vivo by pruning tolerogenic dendritic cells.Cell Rep. 2022 Apr 12;39(2):110657. doi: 10.1016/j.celrep.2022.110657. Cell Rep. 2022. PMID: 35417681 Free PMC article.
-
Epithelial Pyroptosis in Host Defense.J Mol Biol. 2022 Feb 28;434(4):167278. doi: 10.1016/j.jmb.2021.167278. Epub 2021 Oct 8. J Mol Biol. 2022. PMID: 34627788 Free PMC article. Review.
-
PI3KC3 complex subunit NRBF2 is required for apoptotic cell clearance to restrict intestinal inflammation.Autophagy. 2021 May;17(5):1096-1111. doi: 10.1080/15548627.2020.1741332. Epub 2020 Mar 19. Autophagy. 2021. PMID: 32160108 Free PMC article.
-
Inflammasome, Inflammation, and Tissue Homeostasis.Trends Mol Med. 2018 Mar;24(3):304-318. doi: 10.1016/j.molmed.2018.01.004. Epub 2018 Feb 9. Trends Mol Med. 2018. PMID: 29433944 Free PMC article. Review.
-
Macrophage Clearance of Apoptotic Cells: A Critical Assessment.Front Immunol. 2018 Jan 30;9:127. doi: 10.3389/fimmu.2018.00127. eCollection 2018. Front Immunol. 2018. PMID: 29441073 Free PMC article. Review.
References
-
- Bekiaris V, Persson EK, Agace WW. Intestinal dendritic cells in the regulation of mucosal immunity. Immunol Rev. 2014;260:86–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 HD007149/HD/NICHD NIH HHS/United States
- R01 CA161373/CA/NCI NIH HHS/United States
- 5T32DK007792-12/NH/NIH HHS/United States
- U19 AI117873/AI/NIAID NIH HHS/United States
- R01 AI095245/AI/NIAID NIH HHS/United States
- DK072201/NH/NIH HHS/United States
- R01 DK110352/DK/NIDDK NIH HHS/United States
- AI123284/NH/NIH HHS/United States
- R01 DK111862/DK/NIDDK NIH HHS/United States
- HHSN272201000054C/AI/NIAID NIH HHS/United States
- 5R01CA161373-04/NH/NIH HHS/United States
- 5P01DK072201-09/NH/NIH HHS/United States
- T32 DK007792/DK/NIDDK NIH HHS/United States
- U01 DK062429/DK/NIDDK NIH HHS/United States
- U01 DK062422/DK/NIDDK NIH HHS/United States
- P01 DK072201/DK/NIDDK NIH HHS/United States
- AI095245/NH/NIH HHS/United States
- 2T32A1007605-11/NH/NIH HHS/United States
- R01 DK092235/DK/NIDDK NIH HHS/United States
- R01 AI123284/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
