

Phosphorylation of Golgi Peripheral Membrane Protein Grasp65 Is an Integral Step in the Formation of the Human Cytomegalovirus Cytoplasmic Assembly Compartment
- PMID: 27703074
- PMCID: PMC5050342
- DOI: 10.1128/mBio.01554-16
Phosphorylation of Golgi Peripheral Membrane Protein Grasp65 Is an Integral Step in the Formation of the Human Cytomegalovirus Cytoplasmic Assembly Compartment
Abstract
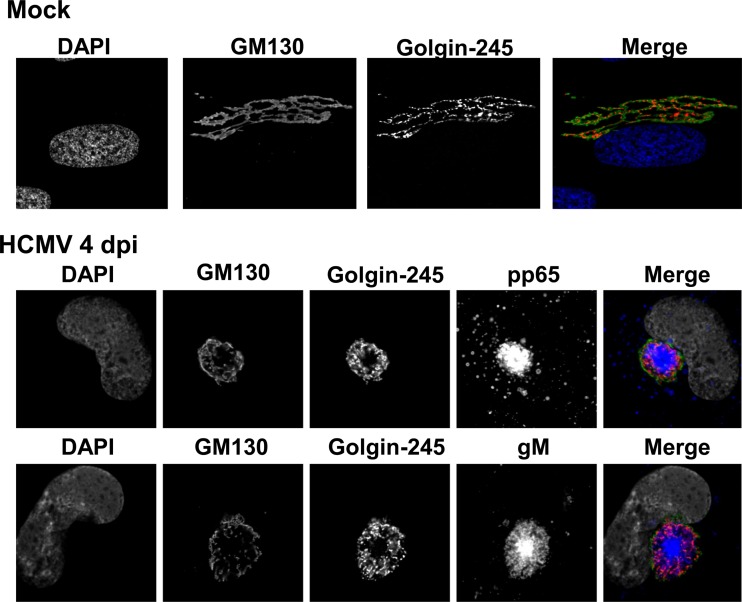
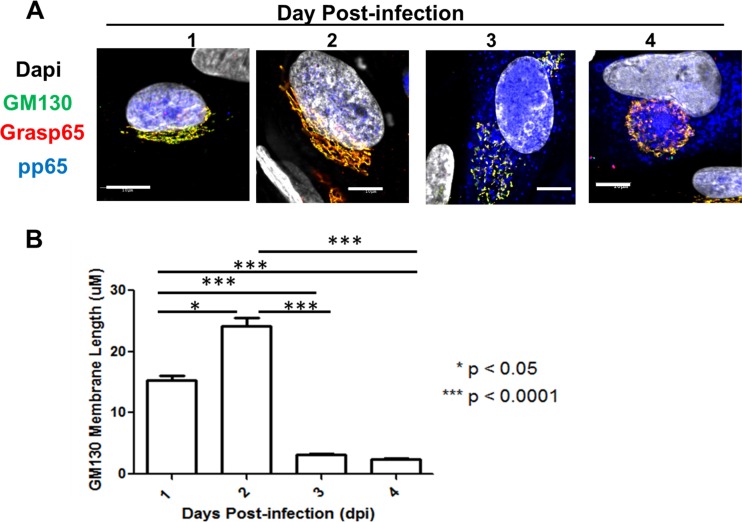
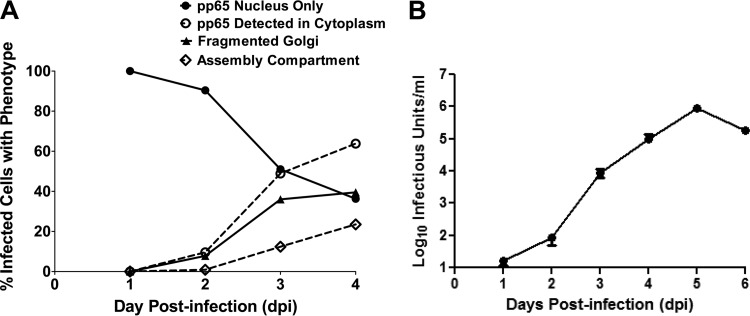
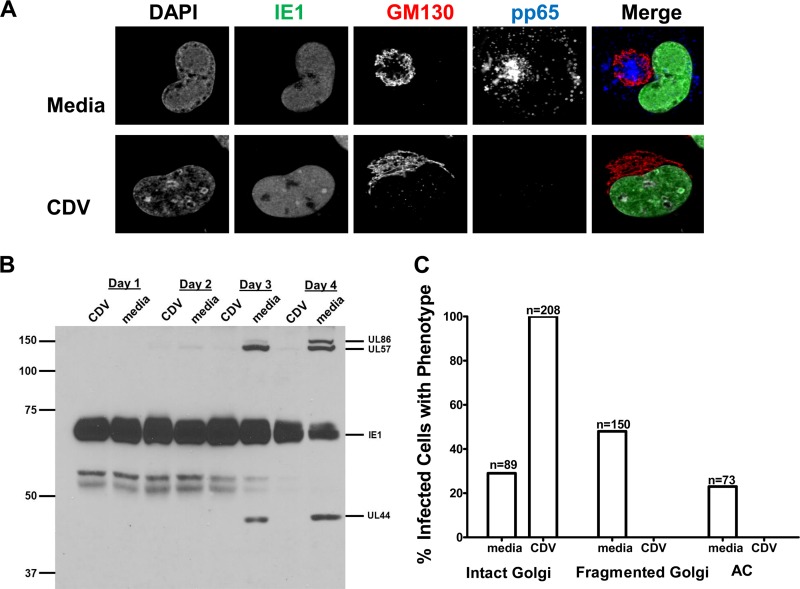
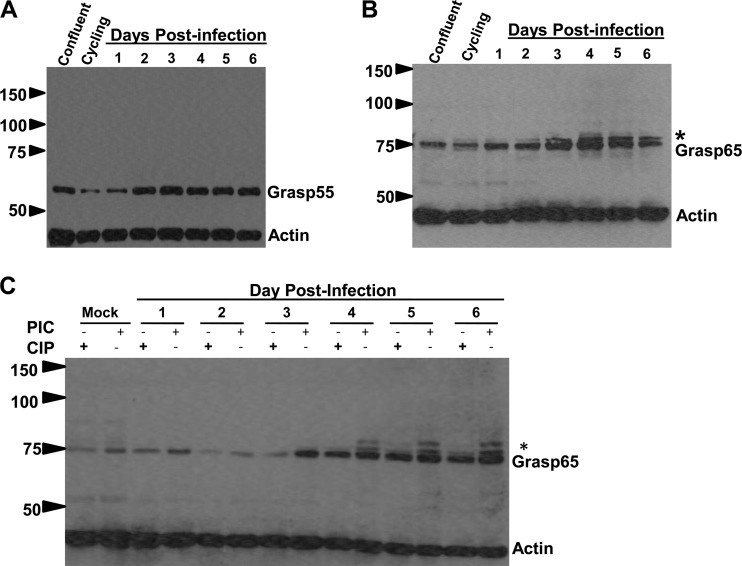
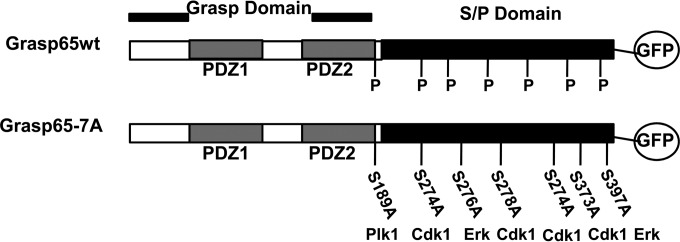
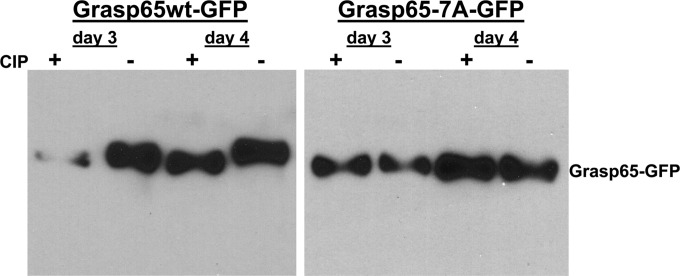
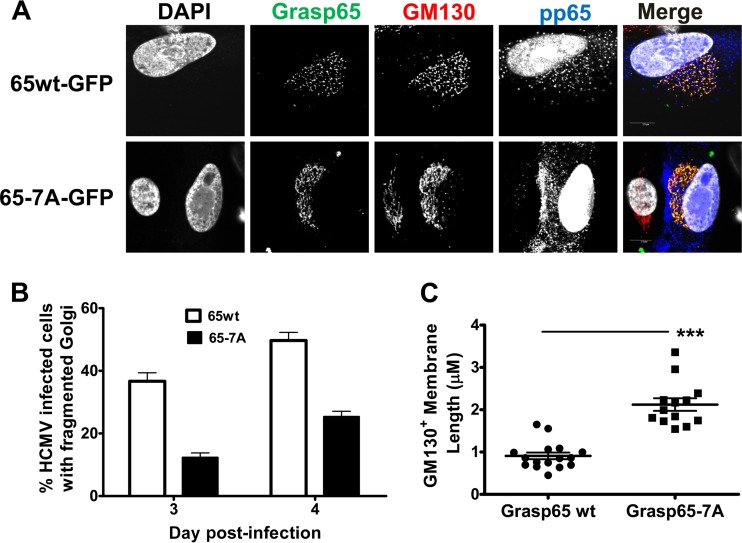
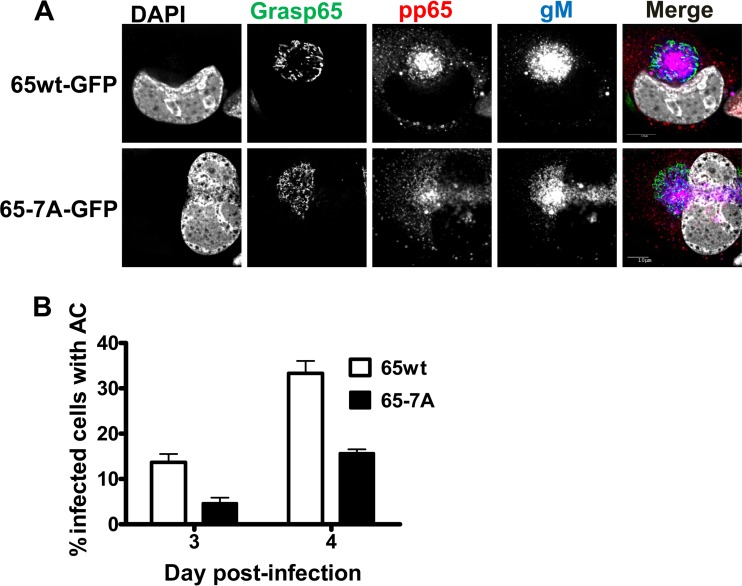
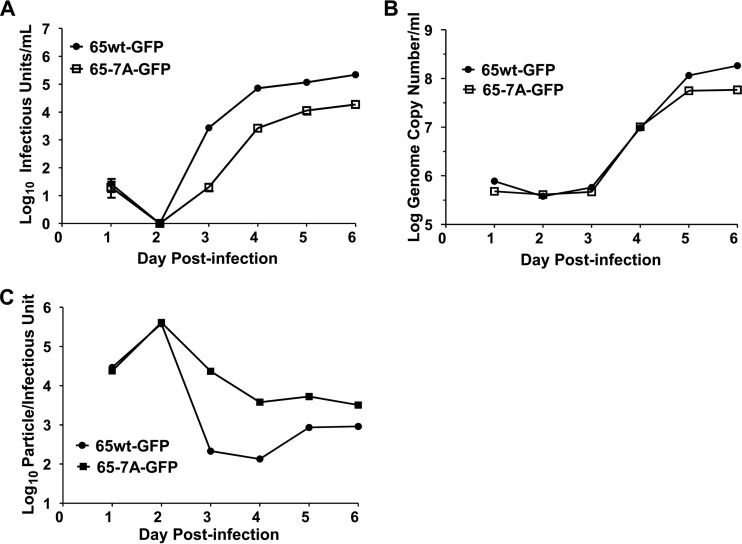
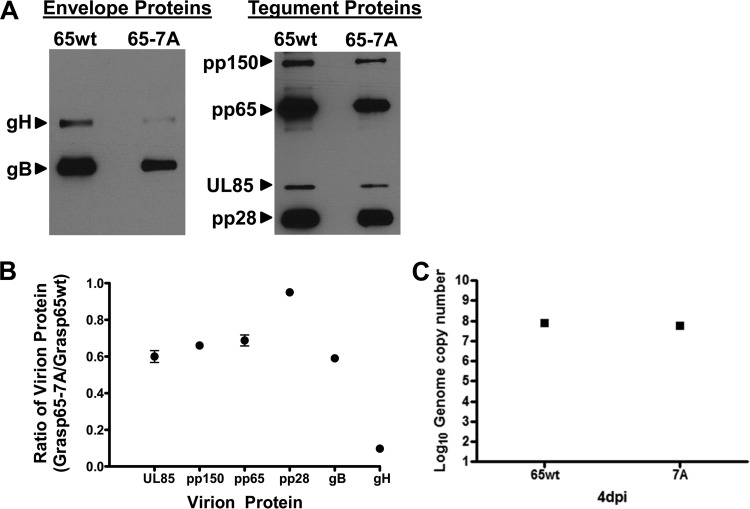
Human cytomegalovirus (HCMV) is the largest member of the Herpesviridae and represents a significant cause of disease. During virus replication, HCMV alters cellular functions to facilitate its replication, including significant reorganization of the secretory and endocytic pathways of the infected cell. A defining morphologic change of the infected cell is the formation of a membranous structure in the cytoplasm that is designated the virion assembly compartment (AC), which consists of virion structural proteins surrounded by cellular membranes. The loss of normal Golgi compartment morphology and its relocalization from a juxtanuclear ribbonlike structure to a series of concentric rings on the periphery of the AC represents a readily recognized reorganization of cellular membranes in the HCMV-infected cell. Although trafficking of viral proteins to this compartment is required for the assembly of infectious virions, the functional significance of the reorganization of intracellular membranes like the Golgi membranes into the AC in the assembly of infectious virus remains understudied. In this study, we determined that Golgi membrane ribbon fragmentation increased during the early cytoplasmic phase of virion assembly and that Golgi membrane fragmentation in infected cells was dependent on the phosphorylation of an integral cis-Golgi protein, Grasp65. Inhibition of Golgi membrane fragmentation and of its reorganization into the AC resulted in decreased production of infectious particles and alteration of the incorporation of an essential protein into the envelope of the mature virion. These results demonstrated the complexity of the virus-host cell interactions required for efficient assembly of this large DNA virus.
Importance: The human cytomegalovirus (HCMV)-induced reorganization of intracellular membranes that is required for the formation of the viral assembly compartment (AC) has been an area of study over the last 20 years. The significance of this virus-induced structure has been evinced by the results of several studies which showed that relocalization of viral proteins to the AC was required for efficient assembly of infectious virus. In this study, we have identified a mechanism for the fragmentation of the Golgi ribbon in the infected cell en route to AC morphogenesis. Identification of this fundamental process during HCMV replication allowed us to propose that the functional role of Golgi membrane reorganization during HCMV infection was the concentration of viral structural proteins and subviral structures into a single intracellular compartment in order to facilitate efficient protein-protein interactions and the virion protein trafficking required for the assembly of this large and structurally complex virus.
Copyright © 2016 Rebmann et al.
Figures
Similar articles
-
Human Cytomegalovirus Envelope Protein gpUL132 Regulates Infectious Virus Production through Formation of the Viral Assembly Compartment.mBio. 2020 Sep 29;11(5):e02044-20. doi: 10.1128/mBio.02044-20. mBio. 2020. PMID: 32994323 Free PMC article.
-
Potent Inhibition of Human Cytomegalovirus by Modulation of Cellular SNARE Syntaxin 5.J Virol. 2016 Dec 16;91(1):e01637-16. doi: 10.1128/JVI.01637-16. Print 2017 Jan 1. J Virol. 2016. PMID: 27795424 Free PMC article.
-
Human Cytomegalovirus Hijacks WD Repeat Domain 11 for Virion Assembly Compartment Formation and Virion Morphogenesis.J Virol. 2022 Mar 9;96(5):e0182721. doi: 10.1128/JVI.01827-21. Epub 2022 Jan 12. J Virol. 2022. PMID: 35020472 Free PMC article.
-
Host Cell Signatures of the Envelopment Site within Beta-Herpes Virions.Int J Mol Sci. 2022 Sep 1;23(17):9994. doi: 10.3390/ijms23179994. Int J Mol Sci. 2022. PMID: 36077391 Free PMC article. Review.
-
Human Cytomegalovirus Egress: Overcoming Barriers and Co-Opting Cellular Functions.Viruses. 2021 Dec 22;14(1):15. doi: 10.3390/v14010015. Viruses. 2021. PMID: 35062219 Free PMC article. Review.
Cited by
-
Human cytomegalovirus deploys molecular mimicry to recruit VPS4A to sites of virus assembly.PLoS Pathog. 2024 Jun 20;20(6):e1012300. doi: 10.1371/journal.ppat.1012300. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38900818 Free PMC article.
-
Human Cytomegalovirus Envelope Protein gpUL132 Regulates Infectious Virus Production through Formation of the Viral Assembly Compartment.mBio. 2020 Sep 29;11(5):e02044-20. doi: 10.1128/mBio.02044-20. mBio. 2020. PMID: 32994323 Free PMC article.
-
The human cytomegalovirus decathlon: Ten critical replication events provide opportunities for restriction.Front Cell Dev Biol. 2022 Nov 25;10:1053139. doi: 10.3389/fcell.2022.1053139. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36506089 Free PMC article. Review.
-
Infection-Induced Changes Within the Endocytic Recycling Compartment Suggest a Roadmap of Human Cytomegalovirus Egress.Front Microbiol. 2018 Aug 22;9:1888. doi: 10.3389/fmicb.2018.01888. eCollection 2018. Front Microbiol. 2018. PMID: 30186245 Free PMC article.
-
Cytomegaloviruses Exploit Recycling Rab Proteins in the Sequential Establishment of the Assembly Compartment.Front Cell Dev Biol. 2018 Dec 4;6:165. doi: 10.3389/fcell.2018.00165. eCollection 2018. Front Cell Dev Biol. 2018. PMID: 30564576 Free PMC article. Review.
References
-
- Yamamoto AY, Mussi-Pinhata MM, Isaac MD, Amaral FR, Carvalheiro CG, Aragon DC, Manfredi AK, Boppana SB, Britt WJ. 2011. Congenital cytomegalovirus infection as a cause of sensorineural hearing loss in a highly immune population. Pediatr Infect Dis J 30:1043–1046. doi:10.1097/INF.0b013e31822d9640. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
