

Tumor macrophages are pivotal constructors of tumor collagenous matrix
- PMID: 27697834
- PMCID: PMC5068227
- DOI: 10.1084/jem.20151193
Tumor macrophages are pivotal constructors of tumor collagenous matrix
Abstract
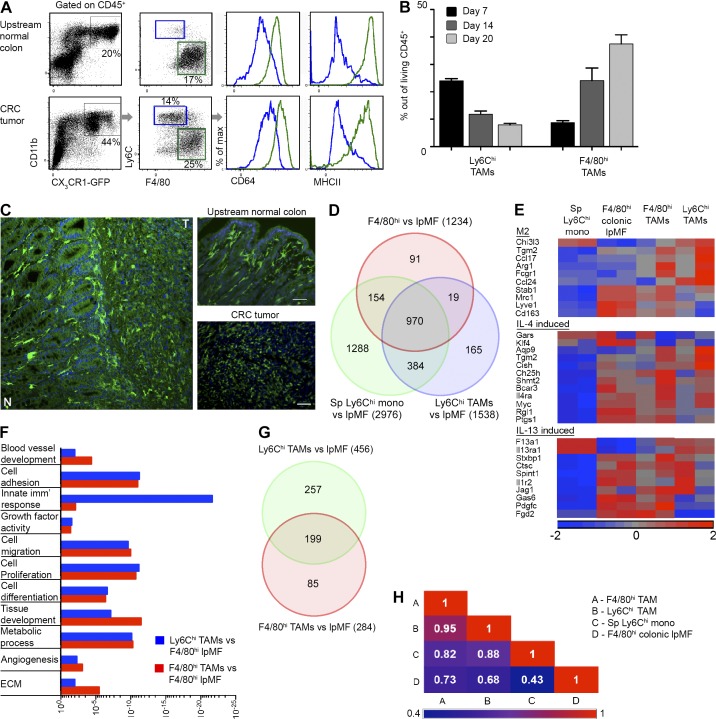
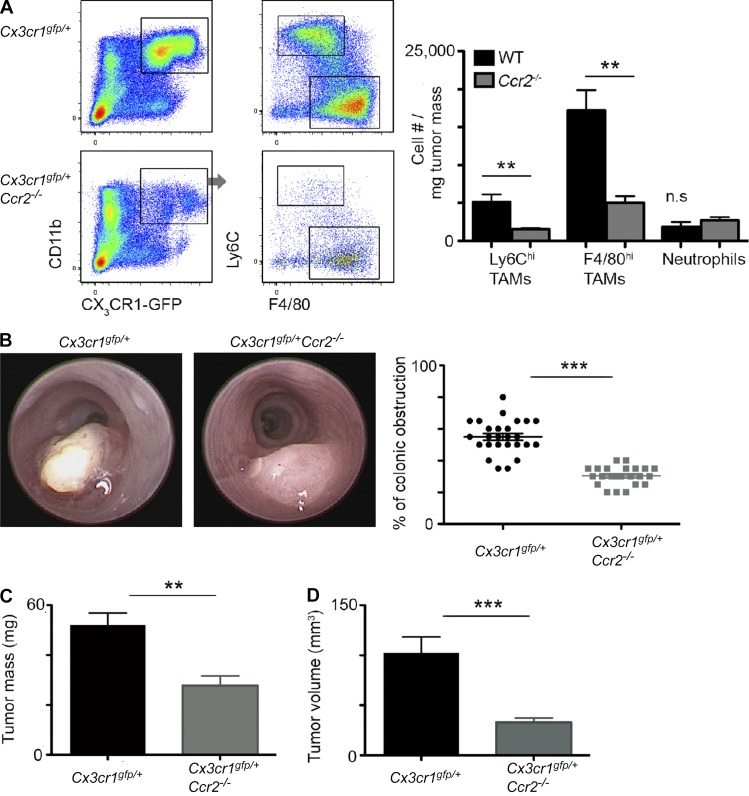
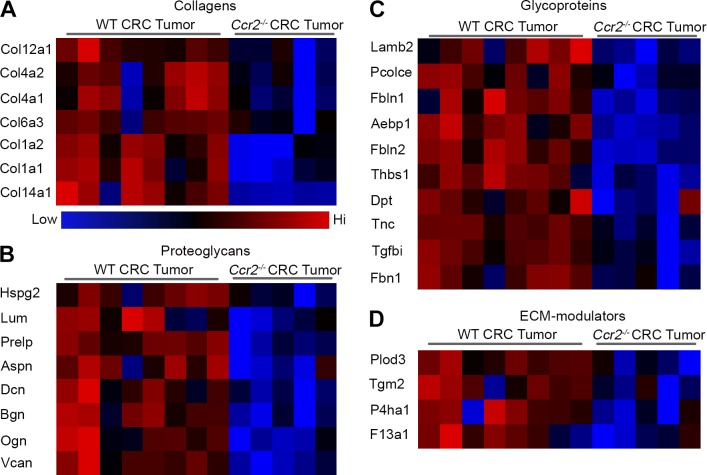
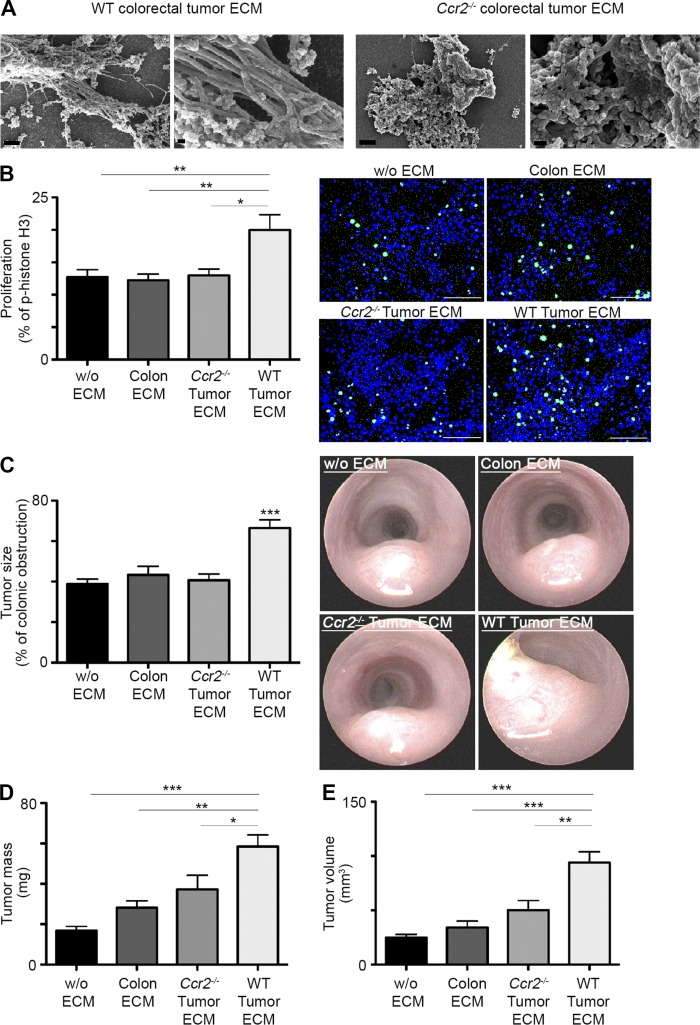
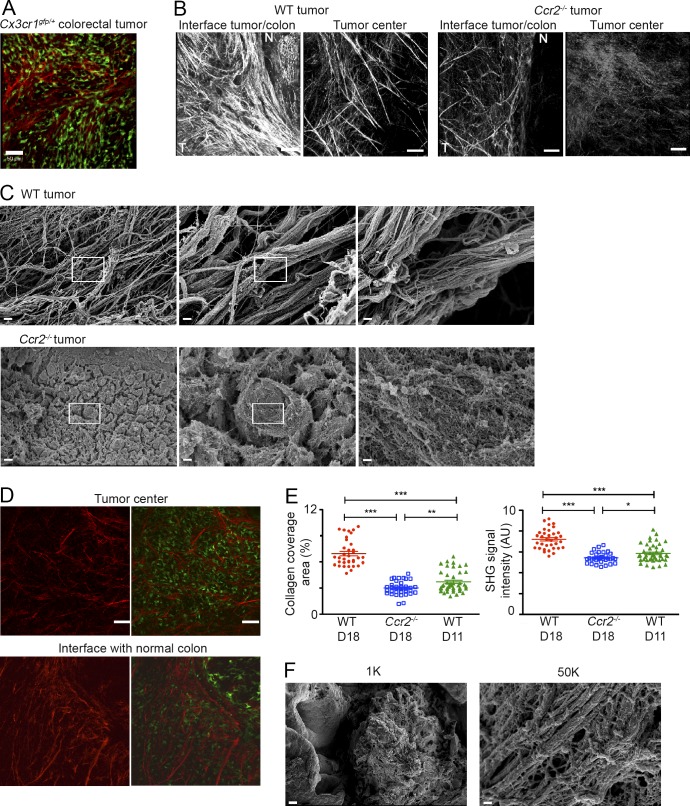
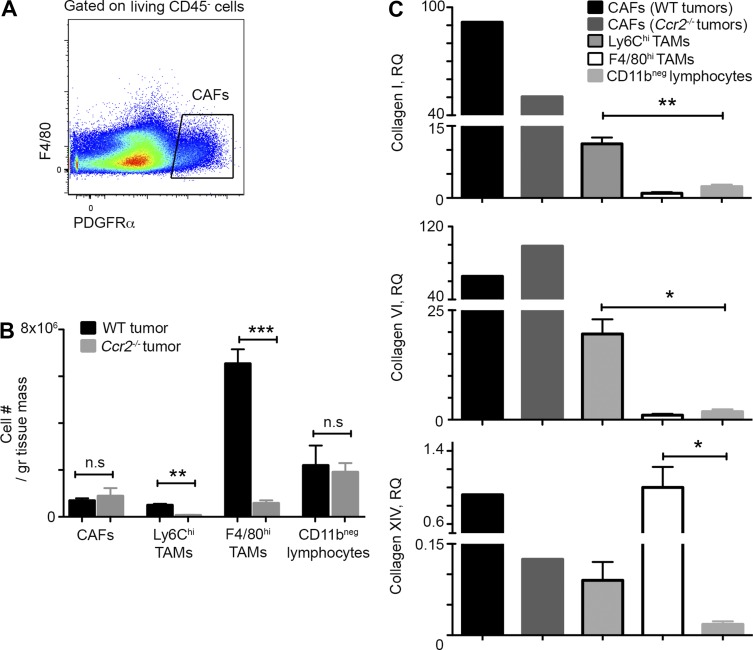
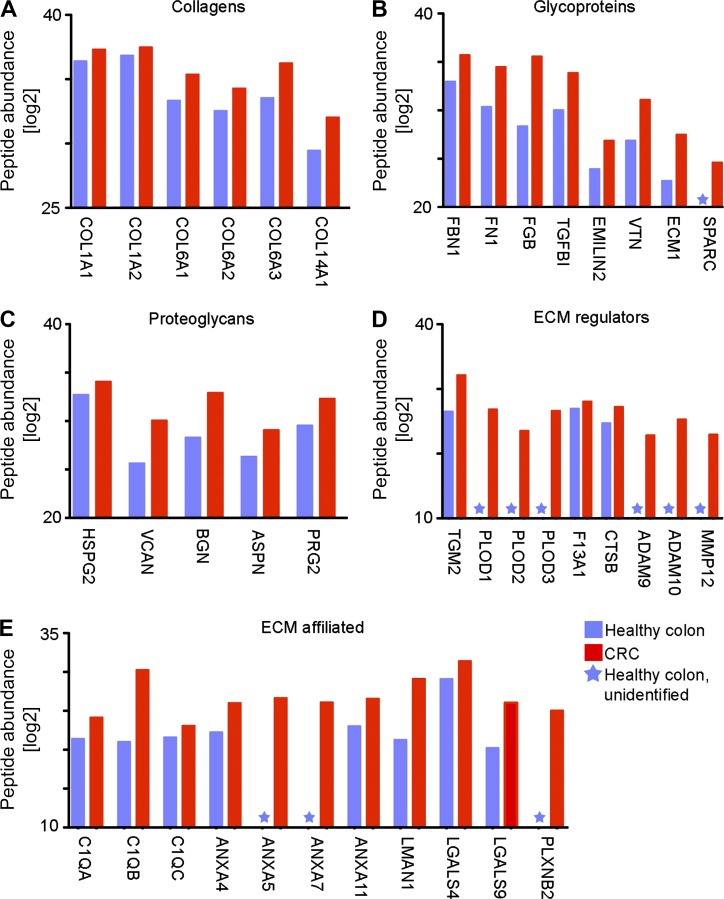
Tumor-associated macrophages (TAMs) promote tumor development, invasion, and dissemination by various mechanisms. In this study, using an orthotopic colorectal cancer (CRC) model, we found that monocyte-derived TAMs advance tumor development by the remodeling of its extracellular matrix (ECM) composition and structure. Unbiased transcriptomic and proteomic analyses of (a) TAM-abundant and -deficient tumor tissues and (b) sorted tumor-associated and -resident colonic macrophage subpopulations defined a distinct TAM-induced ECM molecular signature composed of an ensemble of matricellular proteins and remodeling enzymes they provide to the tumor microenvironment. Remarkably, many of these ECM proteins are specifically increased in human CRC versus healthy colon. Specifically, we demonstrate that although differentiating into TAMs, monocytes up-regulate matrix-remodeling programs associated with the synthesis and assembly of collagenous ECM, specifically collagen types I, VI, and XIV. This finding was further established by advanced imaging showing that TAMs instruct the deposition, cross-linking, and linearization of collagen fibers during tumor development, especially at areas of tumor invasiveness. Finally, we show that cancer-associated fibroblasts are significantly outnumbered by TAMs in this model and that their expression of collagen XIV and I is reduced by TAM deficiency. Here, we outline a novel TAM protumoral function associated with building of the collagenous ECM niche.
© 2016 Afik et al.
Figures

Similar articles
-
Tumorigenic Interplay Between Macrophages and Collagenous Matrix in the Tumor Microenvironment.Methods Mol Biol. 2019;1944:203-220. doi: 10.1007/978-1-4939-9095-5_15. Methods Mol Biol. 2019. PMID: 30840245
-
Stromal fibroblasts shape the myeloid phenotype in normal colon and colorectal cancer and induce CD163 and CCL2 expression in macrophages.Cancer Lett. 2021 Nov 1;520:184-200. doi: 10.1016/j.canlet.2021.07.006. Epub 2021 Jul 10. Cancer Lett. 2021. PMID: 34256095
-
Tumor-Associated Macrophages Derived from Circulating Inflammatory Monocytes Degrade Collagen through Cellular Uptake.Cell Rep. 2017 Dec 26;21(13):3662-3671. doi: 10.1016/j.celrep.2017.12.011. Cell Rep. 2017. PMID: 29281816 Free PMC article.
-
The Role of Tumor-Associated Macrophages in Colorectal Carcinoma Progression.Cell Physiol Biochem. 2018;45(1):356-365. doi: 10.1159/000486816. Epub 2018 Jan 19. Cell Physiol Biochem. 2018. PMID: 29402795 Review.
-
Expression and Function of a Disintegrin and Metalloproteinases in Cancer-Associated Fibroblasts of Colorectal Cancer.Digestion. 2020;101(1):18-24. doi: 10.1159/000504087. Epub 2019 Nov 13. Digestion. 2020. PMID: 31722362 Review.
Cited by
-
Microenvironmental modulation of the developing tumour: an immune-stromal dialogue.Mol Oncol. 2021 Oct;15(10):2600-2633. doi: 10.1002/1878-0261.12773. Epub 2020 Aug 28. Mol Oncol. 2021. PMID: 32741067 Free PMC article. Review.
-
Remodeling the tumor microenvironment via blockade of LAIR-1 and TGF-β signaling enables PD-L1-mediated tumor eradication.J Clin Invest. 2022 Apr 15;132(8):e155148. doi: 10.1172/JCI155148. J Clin Invest. 2022. PMID: 35230974 Free PMC article.
-
The Critical Role of Chemokine (C-C Motif) Receptor 2-Positive Monocytes in Autoimmune Cholangitis.Front Immunol. 2018 Aug 15;9:1852. doi: 10.3389/fimmu.2018.01852. eCollection 2018. Front Immunol. 2018. PMID: 30158929 Free PMC article.
-
Helicobacter pylori-activated fibroblasts as a silent partner in gastric cancer development.Cancer Metastasis Rev. 2023 Dec;42(4):1219-1256. doi: 10.1007/s10555-023-10122-1. Epub 2023 Jul 17. Cancer Metastasis Rev. 2023. PMID: 37460910 Free PMC article. Review.
-
Mitochondrial-related genes markers that predict survival in patients with head and neck squamous cell carcinoma affect immunomodulation through hypoxia, glycolysis, and angiogenesis pathways.Aging (Albany NY). 2023 Oct 4;15(19):10347-10369. doi: 10.18632/aging.205081. Epub 2023 Oct 4. Aging (Albany NY). 2023. PMID: 37796226 Free PMC article.
References
-
- Bain C.C., Scott C.L., Uronen-Hansson H., Gudjonsson S., Jansson O., Grip O., Guilliams M., Malissen B., Agace W.W., and Mowat A.M.. 2013. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal Immunol. 6:498–510. 10.1038/mi.2012.89 - DOI - PMC - PubMed
-
- Bain C.C., Bravo-Blas A., Scott C.L., Gomez Perdiguero E., Geissmann F., Henri S., Malissen B., Osborne L.C., Artis D., and Mowat A.M.. 2014. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15:929–937. 10.1038/ni.2967 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
