

Platelet-derived growth factor-α receptor is the cellular receptor for human cytomegalovirus gHgLgO trimer
- PMID: 27573107
- PMCID: PMC4918640
- DOI: 10.1038/nmicrobiol.2016.82
Platelet-derived growth factor-α receptor is the cellular receptor for human cytomegalovirus gHgLgO trimer
Abstract
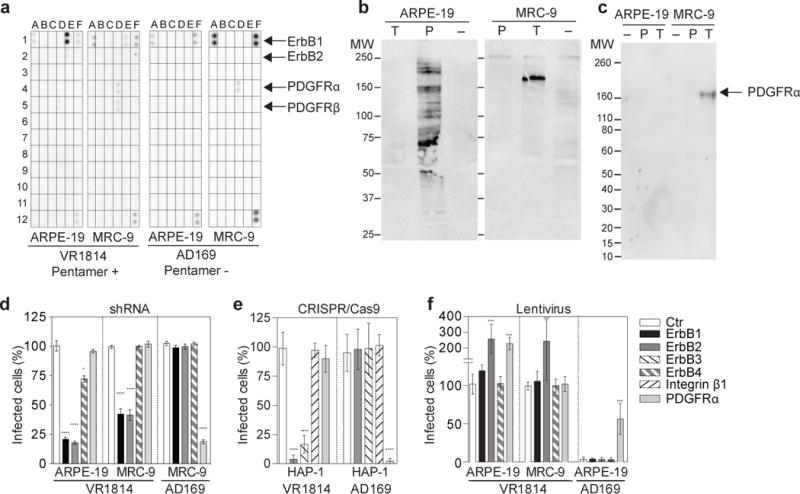
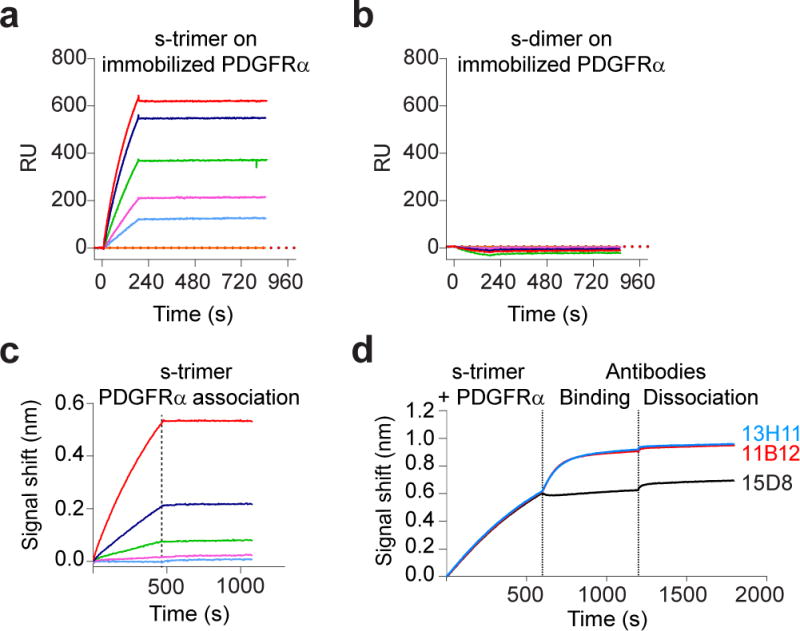
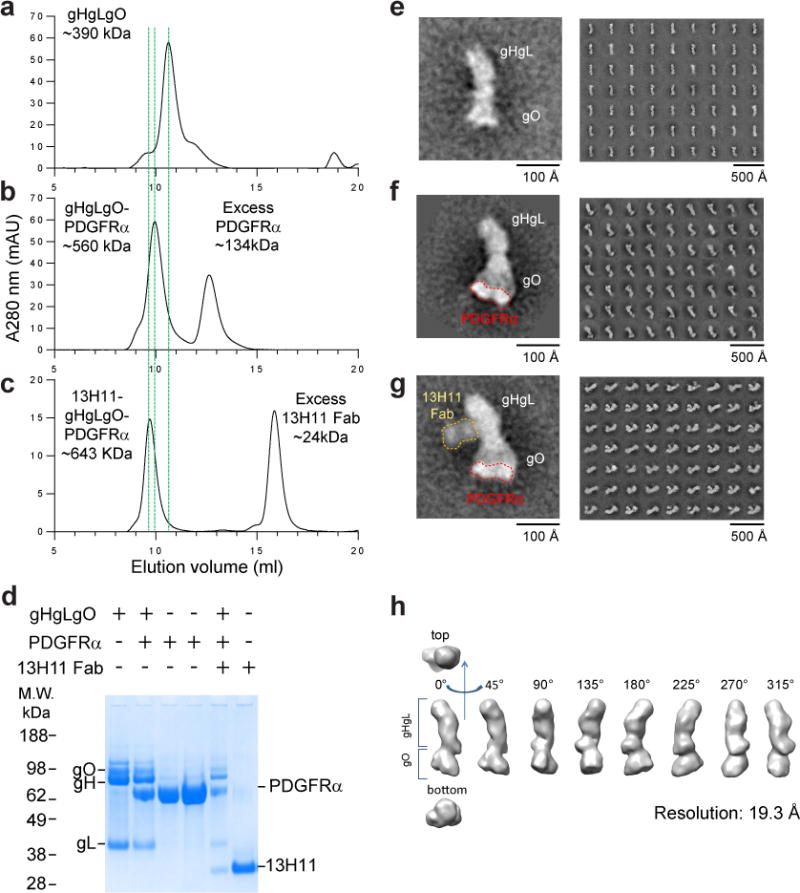
Human cytomegalovirus encodes at least 25 membrane glycoproteins that are found in the viral envelope(1). While gB represents the fusion protein, two glycoprotein complexes control the tropism of the virus: the gHgLgO trimer is involved in the infection of fibroblasts, and the gHgLpUL128L pentamer is required for infection of endothelial, epithelial and myeloid cells(2-5). Two reports suggested that gB binds to ErbB1 and PDGFRα (refs 6,7); however, these results do not explain the tropism of the virus and were recently challenged(8,9). Here, we provide a 19 Å reconstruction for the gHgLgO trimer and show that it binds with high affinity through the gO subunit to PDGFRα, which is expressed on fibroblasts but not on epithelial cells. We also provide evidence that the trimer is essential for viral entry in both fibroblasts and epithelial cells. Furthermore, we identify the pentamer, which is essential for infection of epithelial cells, as a trigger for the ErbB pathway. These findings help explain the broad tropism of human cytomegalovirus and indicate that PDGFRα and the viral gO subunit could be targeted by novel anti-viral therapies.
Conflict of interest statement
Competing interests. The authors declare no financial or commercial conflict of interests.
Figures

Similar articles
-
Cryo-Electron Microscopy Structure and Interactions of the Human Cytomegalovirus gHgLgO Trimer with Platelet-Derived Growth Factor Receptor Alpha.mBio. 2021 Oct 26;12(5):e0262521. doi: 10.1128/mBio.02625-21. Epub 2021 Oct 26. mBio. 2021. PMID: 34700375 Free PMC article.
-
Influence of Human Cytomegalovirus Glycoprotein O Polymorphism on the Inhibitory Effect of Soluble Forms of Trimer- and Pentamer-Specific Entry Receptors.J Virol. 2020 Jul 1;94(14):e00107-20. doi: 10.1128/JVI.00107-20. Print 2020 Jul 1. J Virol. 2020. PMID: 32350071 Free PMC article.
-
Structures of HCMV Trimer reveal the basis for receptor recognition and cell entry.Cell. 2021 Mar 4;184(5):1232-1244.e16. doi: 10.1016/j.cell.2021.01.036. Epub 2021 Feb 23. Cell. 2021. PMID: 33626330
-
Pathogen at the Gates: Human Cytomegalovirus Entry and Cell Tropism.Viruses. 2018 Dec 11;10(12):704. doi: 10.3390/v10120704. Viruses. 2018. PMID: 30544948 Free PMC article. Review.
-
The structural basis of herpesvirus entry.Nat Rev Microbiol. 2021 Feb;19(2):110-121. doi: 10.1038/s41579-020-00448-w. Epub 2020 Oct 21. Nat Rev Microbiol. 2021. PMID: 33087881 Free PMC article. Review.
Cited by
-
Guinea pig cytomegalovirus trimer complex gH/gL/gO uses PDGFRA as universal receptor for cell fusion and entry.Virology. 2020 Sep;548:236-249. doi: 10.1016/j.virol.2020.05.012. Epub 2020 Jun 11. Virology. 2020. PMID: 32791352 Free PMC article.
-
Endoplasmic Reticulum (ER) Reorganization and Intracellular Retention of CD58 Are Functionally Independent Properties of the Human Cytomegalovirus ER-Resident Glycoprotein UL148.J Virol. 2020 Feb 14;94(5):e01435-19. doi: 10.1128/JVI.01435-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31801856 Free PMC article.
-
The Human Cytomegalovirus Nonstructural Glycoprotein UL148 Reorganizes the Endoplasmic Reticulum.mBio. 2019 Dec 10;10(6):e02110-19. doi: 10.1128/mBio.02110-19. mBio. 2019. PMID: 31822584 Free PMC article.
-
Expression Levels of Glycoprotein O (gO) Vary between Strains of Human Cytomegalovirus, Influencing the Assembly of gH/gL Complexes and Virion Infectivity.J Virol. 2018 Jul 17;92(15):e00606-18. doi: 10.1128/JVI.00606-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29743375 Free PMC article.
-
Selection of Human Cytomegalovirus Mutants with Resistance against PDGFRα-Derived Entry Inhibitors.Viruses. 2021 Jun 8;13(6):1094. doi: 10.3390/v13061094. Viruses. 2021. PMID: 34201364 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
