

Role of Nfu1 and Bol3 in iron-sulfur cluster transfer to mitochondrial clients
- PMID: 27532773
- PMCID: PMC5014551
- DOI: 10.7554/eLife.15991
Role of Nfu1 and Bol3 in iron-sulfur cluster transfer to mitochondrial clients
Abstract
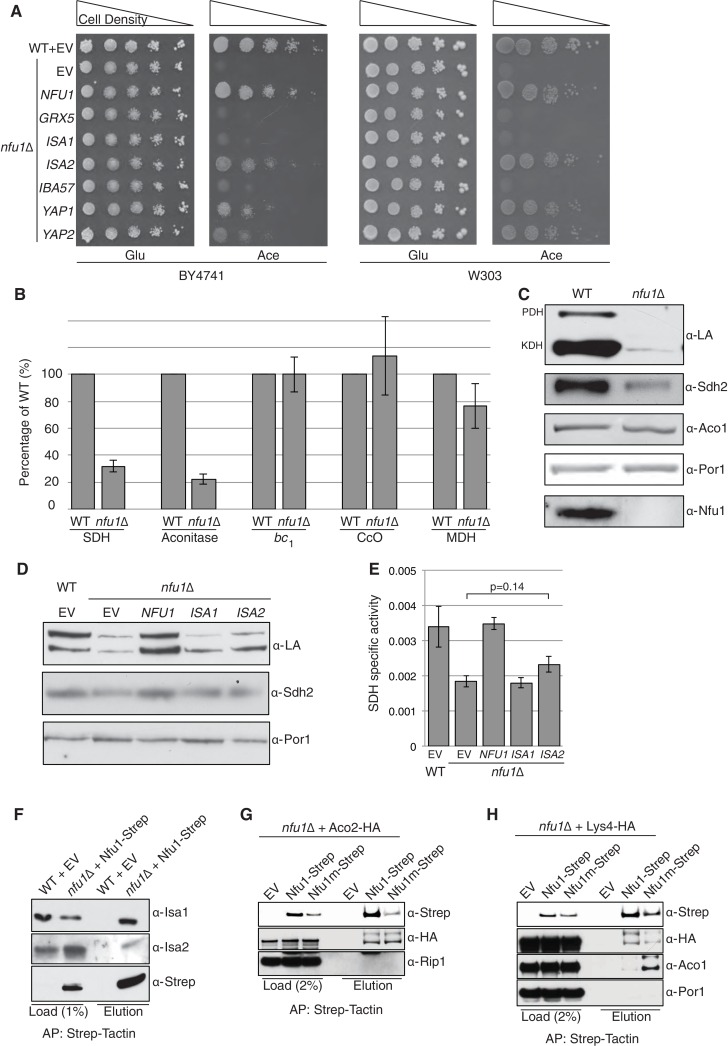
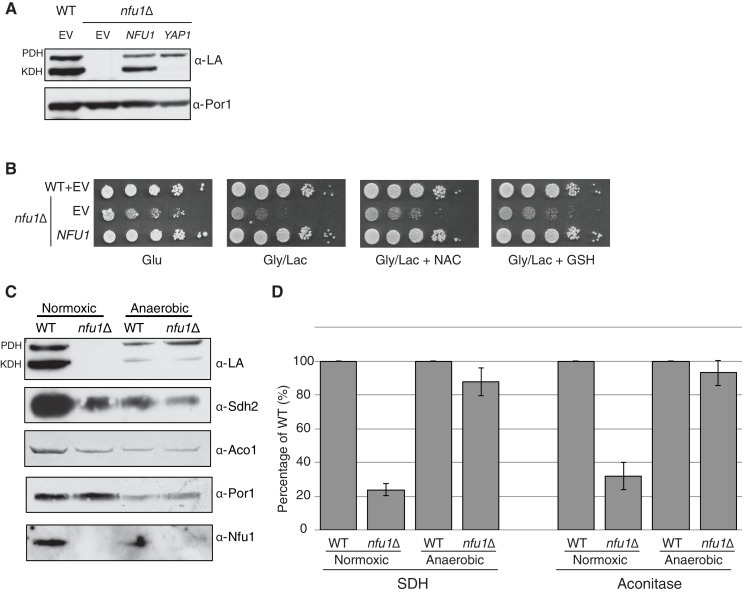
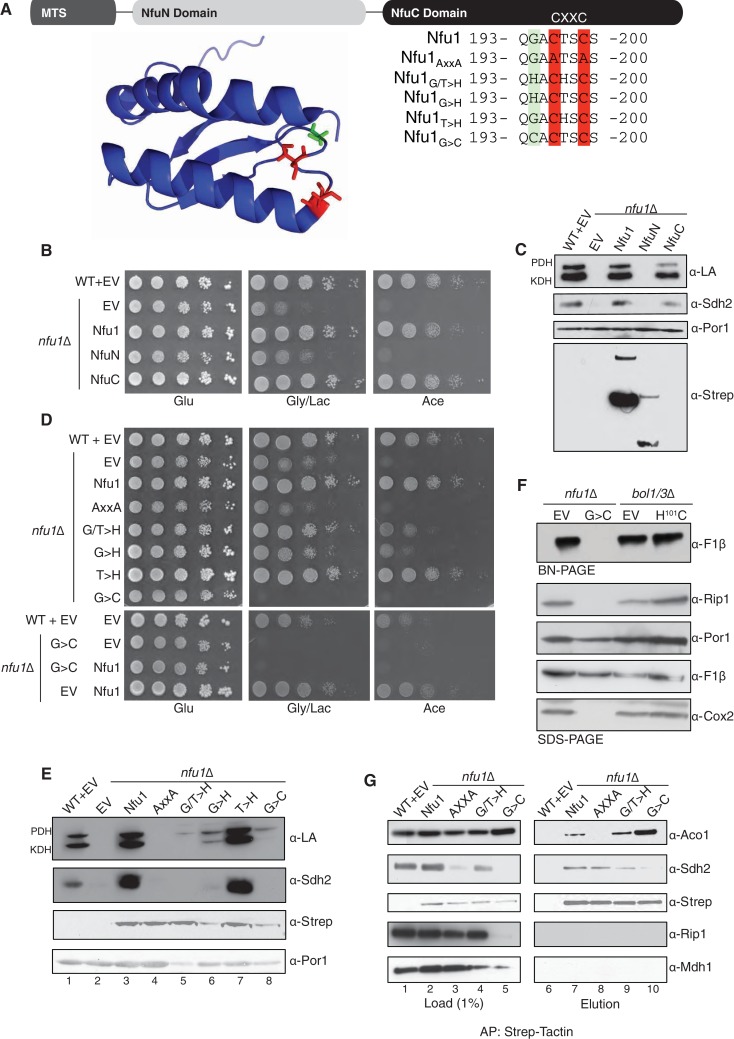
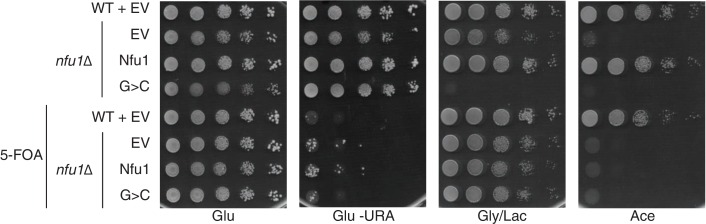
Iron-sulfur (Fe-S) clusters are essential for many cellular processes, ranging from aerobic respiration, metabolite biosynthesis, ribosome assembly and DNA repair. Mutations in NFU1 and BOLA3 have been linked to genetic diseases with defects in mitochondrial Fe-S centers. Through genetic studies in yeast, we demonstrate that Nfu1 functions in a late step of [4Fe-4S] cluster biogenesis that is of heightened importance during oxidative metabolism. Proteomic studies revealed Nfu1 physical interacts with components of the ISA [4Fe-4S] assembly complex and client proteins that need [4Fe-4S] clusters to function. Additional studies focused on the mitochondrial BolA proteins, Bol1 and Bol3 (yeast homolog to human BOLA3), revealing that Bol1 functions earlier in Fe-S biogenesis with the monothiol glutaredoxin, Grx5, and Bol3 functions late with Nfu1. Given these observations, we propose that Nfu1, assisted by Bol3, functions to facilitate Fe-S transfer from the biosynthetic apparatus to the client proteins preventing oxidative damage to [4Fe-4S] clusters.
Keywords: S. cerevisiae; biochemistry; cofactor biogenesis; iron-sulfur biogenesis; iron-sulfur clusters; mitochondria; oxidative damage.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures


Similar articles
-
Mitochondrial Bol1 and Bol3 function as assembly factors for specific iron-sulfur proteins.Elife. 2016 Aug 17;5:e16673. doi: 10.7554/eLife.16673. Elife. 2016. PMID: 27532772 Free PMC article.
-
The iron-sulfur cluster assembly (ISC) protein Iba57 executes a tetrahydrofolate-independent function in mitochondrial [4Fe-4S] protein maturation.J Biol Chem. 2022 Oct;298(10):102465. doi: 10.1016/j.jbc.2022.102465. Epub 2022 Sep 6. J Biol Chem. 2022. PMID: 36075292 Free PMC article.
-
A pathway for assembling [4Fe-4S]2+ clusters in mitochondrial iron-sulfur protein biogenesis.FEBS J. 2020 Jun;287(11):2312-2327. doi: 10.1111/febs.15140. Epub 2019 Dec 3. FEBS J. 2020. PMID: 31724821
-
Steps Toward Understanding Mitochondrial Fe/S Cluster Biogenesis.Methods Enzymol. 2018;599:265-292. doi: 10.1016/bs.mie.2017.09.004. Epub 2017 Nov 27. Methods Enzymol. 2018. PMID: 29746243 Review.
-
Molecular Basis of Rare Diseases Associated to the Maturation of Mitochondrial [4Fe-4S]-Containing Proteins.Biomolecules. 2022 Jul 21;12(7):1009. doi: 10.3390/biom12071009. Biomolecules. 2022. PMID: 35883565 Free PMC article. Review.
Cited by
-
Iron-sulfur cluster biogenesis, trafficking, and signaling: Roles for CGFS glutaredoxins and BolA proteins.Biochim Biophys Acta Mol Cell Res. 2021 Jan;1868(1):118847. doi: 10.1016/j.bbamcr.2020.118847. Epub 2020 Sep 7. Biochim Biophys Acta Mol Cell Res. 2021. PMID: 32910989 Free PMC article. Review.
-
Roles and maturation of iron-sulfur proteins in plastids.J Biol Inorg Chem. 2018 Jun;23(4):545-566. doi: 10.1007/s00775-018-1532-1. Epub 2018 Jan 18. J Biol Inorg Chem. 2018. PMID: 29349662 Free PMC article. Review.
-
A biochemical sulfur delivery service.Science. 2017 Oct 20;358(6361):307-308. doi: 10.1126/science.aap9299. Science. 2017. PMID: 29051366 Free PMC article. No abstract available.
-
[4Fe-4S] cluster trafficking mediated by Arabidopsis mitochondrial ISCA and NFU proteins.J Biol Chem. 2020 Dec 25;295(52):18367-18378. doi: 10.1074/jbc.RA120.015726. Epub 2020 Oct 29. J Biol Chem. 2020. PMID: 33122194 Free PMC article.
-
Cytosolic iron-sulfur cluster transfer-a proposed kinetic pathway for reconstitution of glutaredoxin 3.FEBS Lett. 2016 Dec;590(24):4531-4540. doi: 10.1002/1873-3468.12491. Epub 2016 Dec 1. FEBS Lett. 2016. PMID: 27859051 Free PMC article.
References
-
- Al-Hassnan ZN, Al-Dosary M, Alfadhel M, Faqeih EA, Alsagob M, Kenana R, Almass R, Al-Harazi OS, Al-Hindi H, Malibari OI, Almutari FB, Tulbah S, Alhadeq F, Al-Sheddi T, Alamro R, AlAsmari A, Almuntashri M, Alshaalan H, Al-Mohanna FA, Colak D, Kaya N. ISCA2 mutation causes infantile neurodegenerative mitochondrial disorder. Journal of Medical Genetics. 2015;52:186–194. doi: 10.1136/jmedgenet-2014-102592. - DOI - PubMed
-
- Angelini S, Gerez C, Ollagnier-de Choudens S, Sanakis Y, Fontecave M, Barras F, Py B. NfuA, a new factor required for maturing Fe/S proteins in Escherichia coli under oxidative stress and iron starvation conditions. Journal of Biological Chemistry. 2008;283:14084–14091. doi: 10.1074/jbc.M709405200. - DOI - PubMed
-
- Baker PR, Friederich MW, Swanson MA, Shaikh T, Bhattacharya K, Scharer GH, Aicher J, Creadon-Swindell G, Geiger E, MacLean KN, Lee WT, Deshpande C, Freckmann ML, Shih LY, Wasserstein M, Rasmussen MB, Lund AM, Procopis P, Cameron JM, Robinson BH, Brown GK, Brown RM, Compton AG, Dieckmann CL, Collard R, Coughlin CR, Spector E, Wempe MF, Van Hove JL. Variant non ketotic hyperglycinemia is caused by mutations in LIAS, BOLA3 and the novel gene GLRX5. Brain. 2014;137:366–379. doi: 10.1093/brain/awt328. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
