

B Virus (Macacine Herpesvirus 1) Divergence: Variations in Glycoprotein D from Clinical and Laboratory Isolates Diversify Virus Entry Strategies
- PMID: 27512063
- PMCID: PMC5044838
- DOI: 10.1128/JVI.00799-16
B Virus (Macacine Herpesvirus 1) Divergence: Variations in Glycoprotein D from Clinical and Laboratory Isolates Diversify Virus Entry Strategies
Abstract
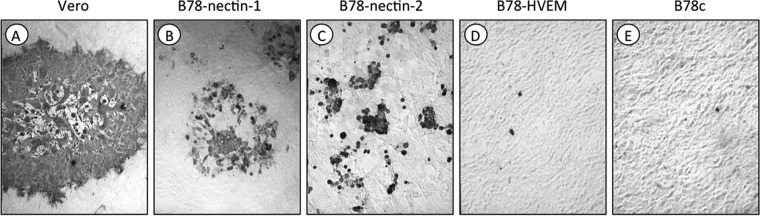
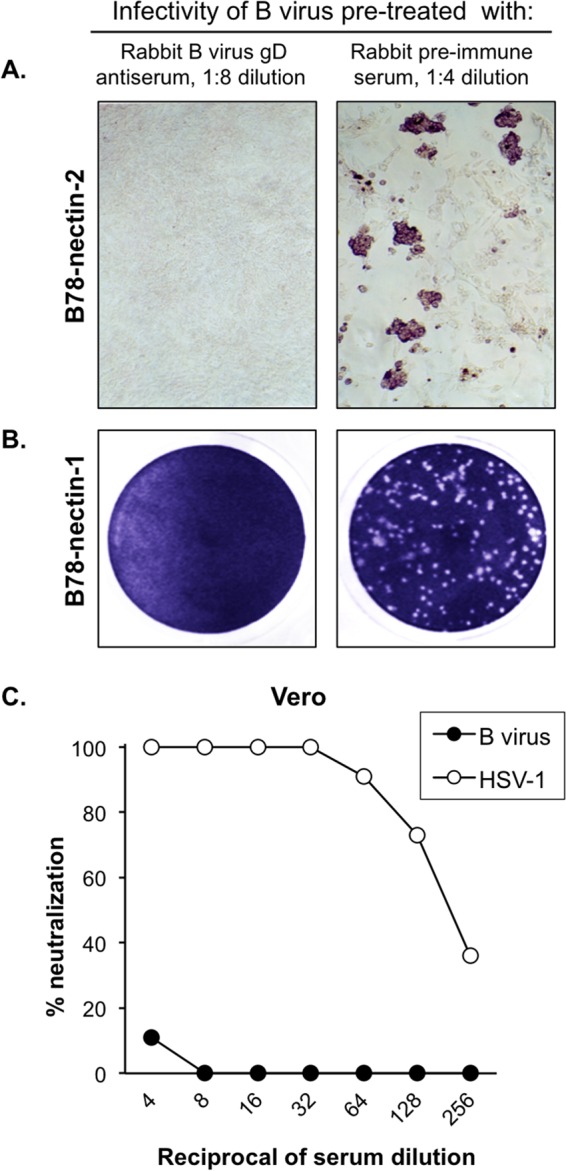
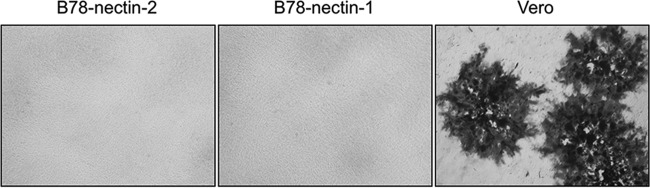
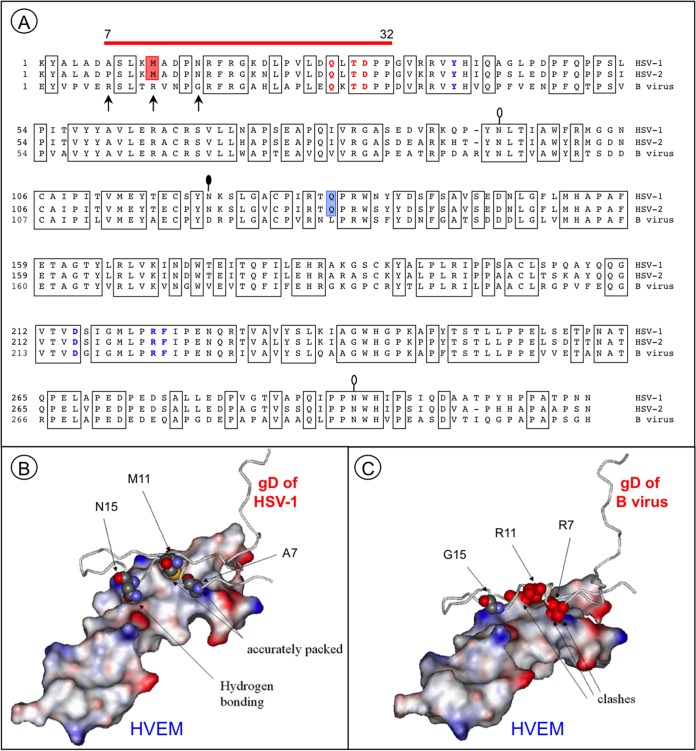
B virus (Macacine herpesvirus 1) can cause deadly zoonotic disease in humans. Molecular mechanisms of B virus cell entry are poorly understood for both macaques and humans. Here we investigated the abilities of clinical B virus isolates to use entry receptors of herpes simplex viruses (HSV). We showed that resistant B78H1 cells became susceptible to B virus clinical strains upon expression of either human nectin-2 or nectin-1. Antibody against glycoprotein D (gD) protected these nectin-bearing cells from B virus infection, and a gD-negative recombinant B virus failed to enter these cells, indicating that the nectin-mediated B virus entry depends on gD. We observed that the infectivity of B virus isolates with a single amino acid substitution (D122N) in the IgV-core of the gD ectodomain was impaired on nectin-1-bearing cells. Computational homology-based modeling of the B virus gD-nectin-1 complex revealed conformational differences between the structures of the gD-122N and gD-122D variants that affected the gD-nectin-1 protein-protein interface and binding affinity. Unlike HSV, B virus clinical strains were unable to use herpesvirus entry mediator (HVEM) as a receptor, regardless of conservation of the gD amino acid residues essential for HSV-1 entry via HVEM. Based on the model of the B virus gD-HVEM interface, we predict that residues R7, R11, and G15 are largely responsible for the inability of B virus to utilize HVEM for entry. The ability of B virus to enter cells of a human host by using a combination of receptors distinct from those for HSV-1 or HSV-2 suggests a possible mechanism of enhanced neuropathogenicity associated with zoonotic infections.
Importance: B virus causes brainstem destruction in infected humans in the absence of timely diagnosis and intervention. Nectins are cell adhesion molecules that are widely expressed in human tissues, including neurons and neuronal synapses. Here we report that human nectin-2 is a target receptor for B virus entry, in addition to the reported receptor human nectin-1. Similar to a B virus lab strain, B virus clinical strains can effectively use both nectin-1 and nectin-2 as cellular receptors for entry into human cells, but unlike HSV-1 and HSV-2, none of the clinical strains uses an HVEM-mediated entry pathway. Ultimately, these differences between B virus and HSV-1 and -2 may provide insight into the neuropathogenicity of B virus during zoonotic infections.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Mutations in the N termini of herpes simplex virus type 1 and 2 gDs alter functional interactions with the entry/fusion receptors HVEM, nectin-2, and 3-O-sulfated heparan sulfate but not with nectin-1.J Virol. 2003 Sep;77(17):9221-31. doi: 10.1128/jvi.77.17.9221-9231.2003. J Virol. 2003. PMID: 12915538 Free PMC article.
-
Herpes B virus utilizes human nectin-1 but not HVEM or PILRα for cell-cell fusion and virus entry.J Virol. 2012 Apr;86(8):4468-76. doi: 10.1128/JVI.00041-12. Epub 2012 Feb 15. J Virol. 2012. PMID: 22345445 Free PMC article.
-
Mutations in herpes simplex virus glycoprotein D that prevent cell entry via nectins and alter cell tropism.Proc Natl Acad Sci U S A. 2004 Aug 24;101(34):12414-21. doi: 10.1073/pnas.0404211101. Epub 2004 Jul 23. Proc Natl Acad Sci U S A. 2004. PMID: 15273289 Free PMC article.
-
Herpesvirus Entry Mediator and Ocular Herpesvirus Infection: More than Meets the Eye.J Virol. 2017 Jun 9;91(13):e00115-17. doi: 10.1128/JVI.00115-17. Print 2017 Jul 1. J Virol. 2017. PMID: 28404853 Free PMC article. Review.
-
The structural basis of herpesvirus entry.Nat Rev Microbiol. 2021 Feb;19(2):110-121. doi: 10.1038/s41579-020-00448-w. Epub 2020 Oct 21. Nat Rev Microbiol. 2021. PMID: 33087881 Free PMC article. Review.
Cited by
-
Archaic connectivity between the sulfated heparan sulfate and the herpesviruses - An evolutionary potential for cross-species interactions.Comput Struct Biotechnol J. 2023 Jan 13;21:1030-1040. doi: 10.1016/j.csbj.2023.01.005. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 36733705 Free PMC article. Review.
-
Development of a vesicular stomatitis virus pseudotyped with herpes B virus glycoproteins and its application in a neutralizing antibody detection assay.mBio. 2024 Jul 17;15(7):e0109224. doi: 10.1128/mbio.01092-24. Epub 2024 Jun 7. mBio. 2024. PMID: 38847539 Free PMC article.
-
Infectious vaccine-derived rubella viruses emerge, persist, and evolve in cutaneous granulomas of children with primary immunodeficiencies.PLoS Pathog. 2019 Oct 28;15(10):e1008080. doi: 10.1371/journal.ppat.1008080. eCollection 2019 Oct. PLoS Pathog. 2019. PMID: 31658304 Free PMC article.
-
Primate Simplexviruses Differ in Tropism for Macaque Cells.Microorganisms. 2022 Dec 21;11(1):26. doi: 10.3390/microorganisms11010026. Microorganisms. 2022. PMID: 36677317 Free PMC article.
-
Identification of unique B virus (Macacine Herpesvirus 1) epitopes of zoonotic and macaque isolates using monoclonal antibodies.PLoS One. 2017 Aug 4;12(8):e0182355. doi: 10.1371/journal.pone.0182355. eCollection 2017. PLoS One. 2017. PMID: 28783746 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
