

Nucleolin-Mediated RNA Localization Regulates Neuron Growth and Cycling Cell Size
- PMID: 27477284
- PMCID: PMC4978702
- DOI: 10.1016/j.celrep.2016.07.005
Nucleolin-Mediated RNA Localization Regulates Neuron Growth and Cycling Cell Size
Abstract
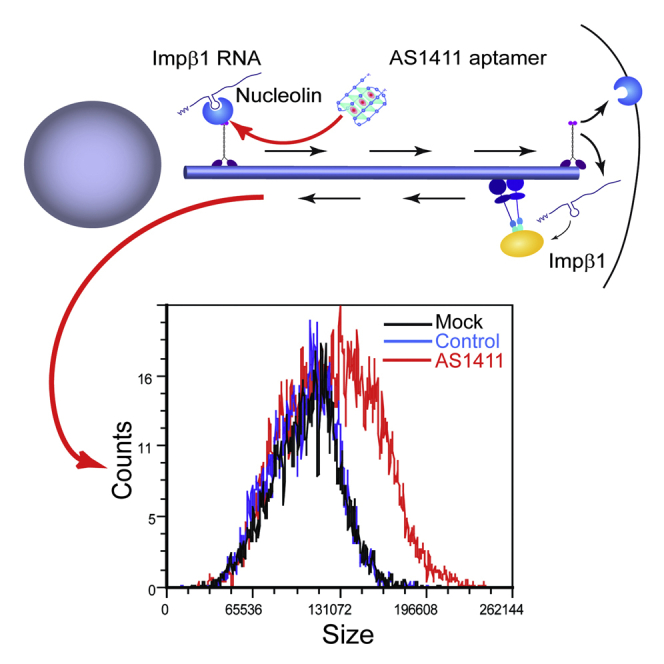
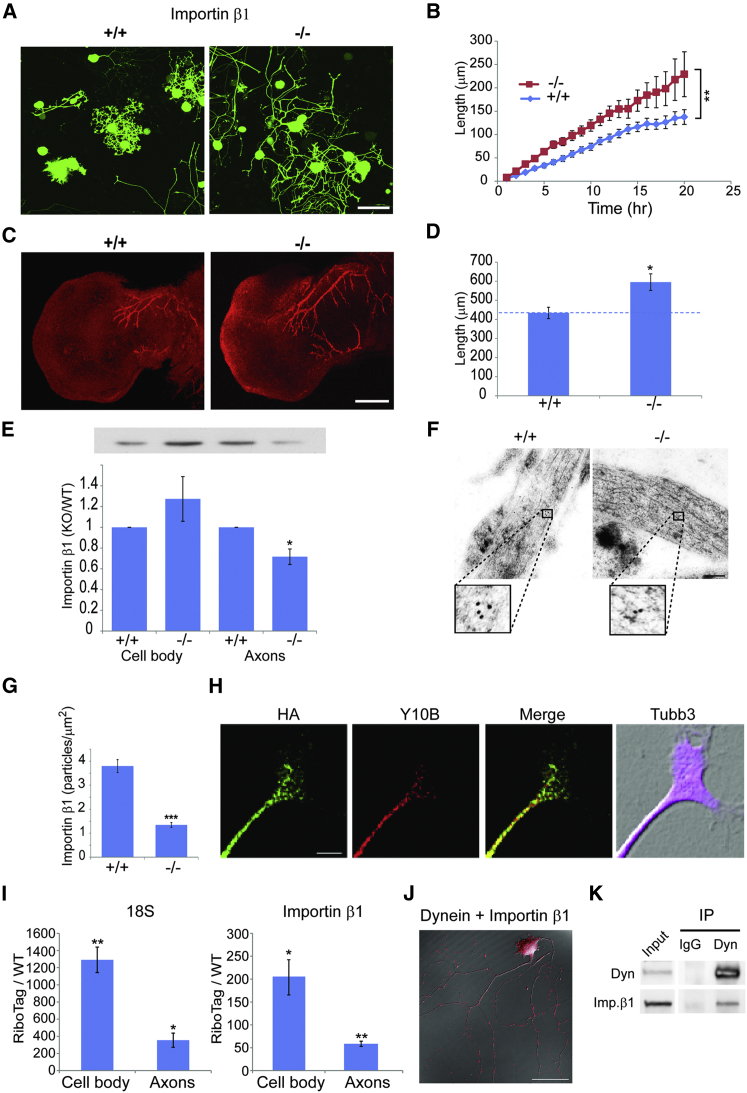
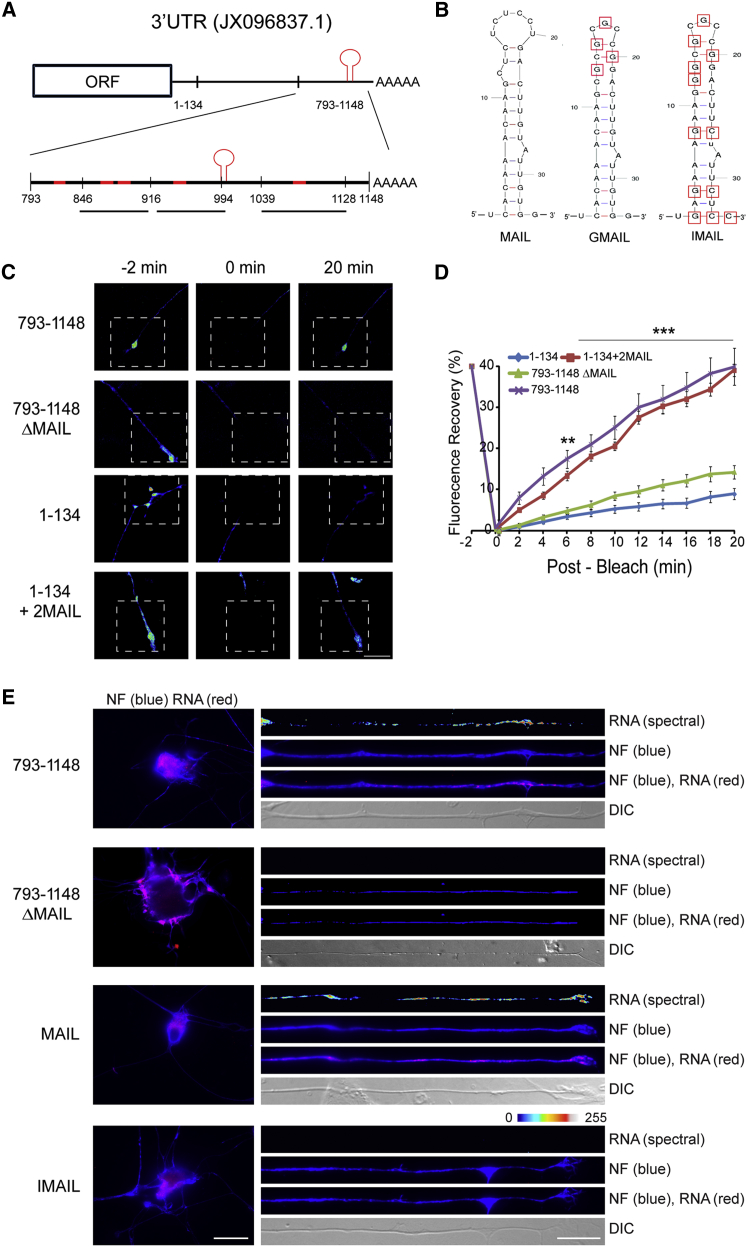
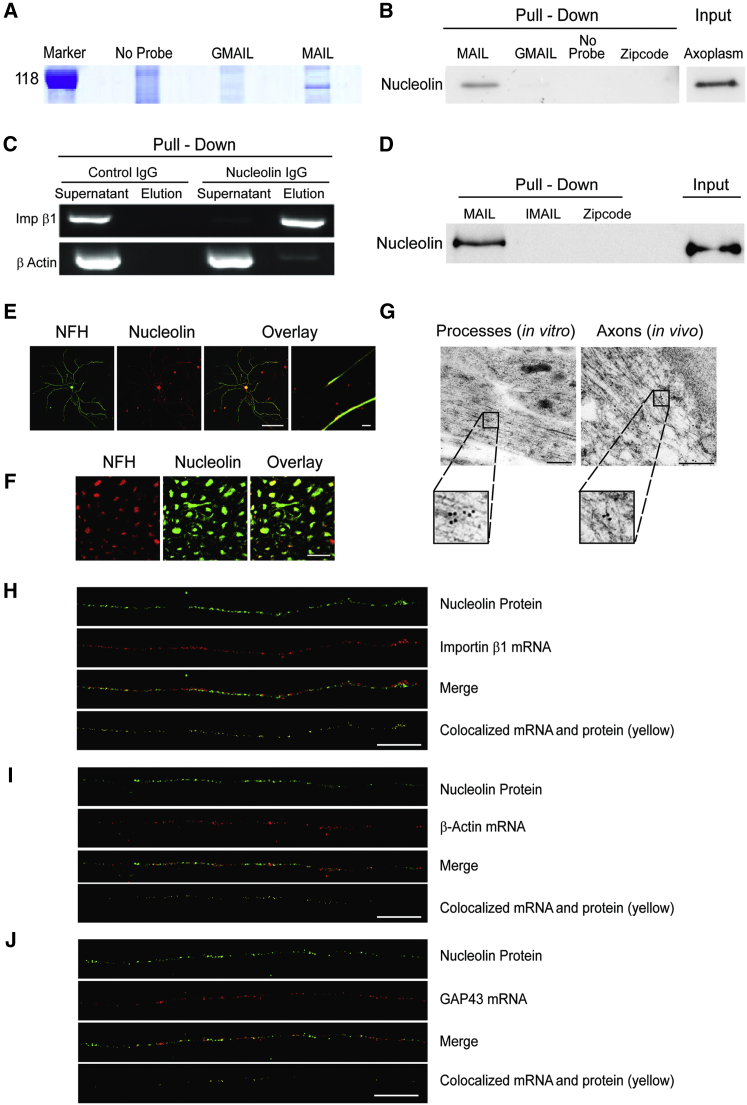
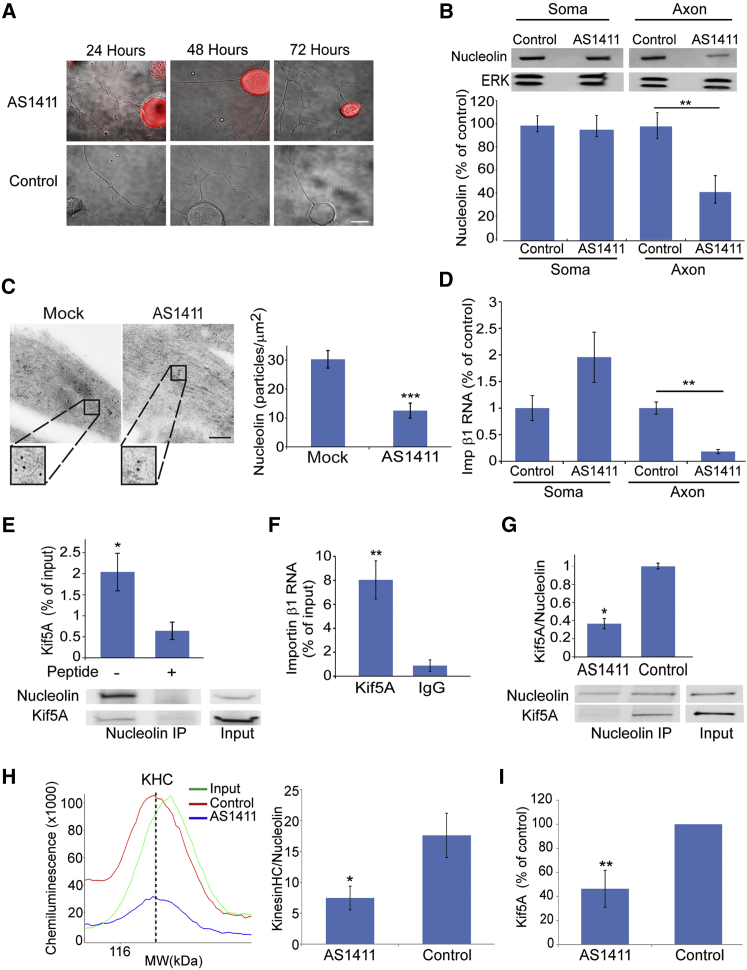
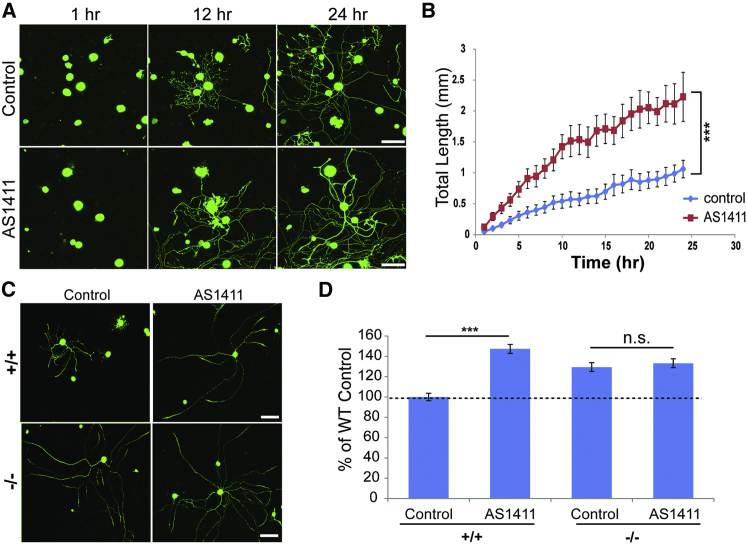
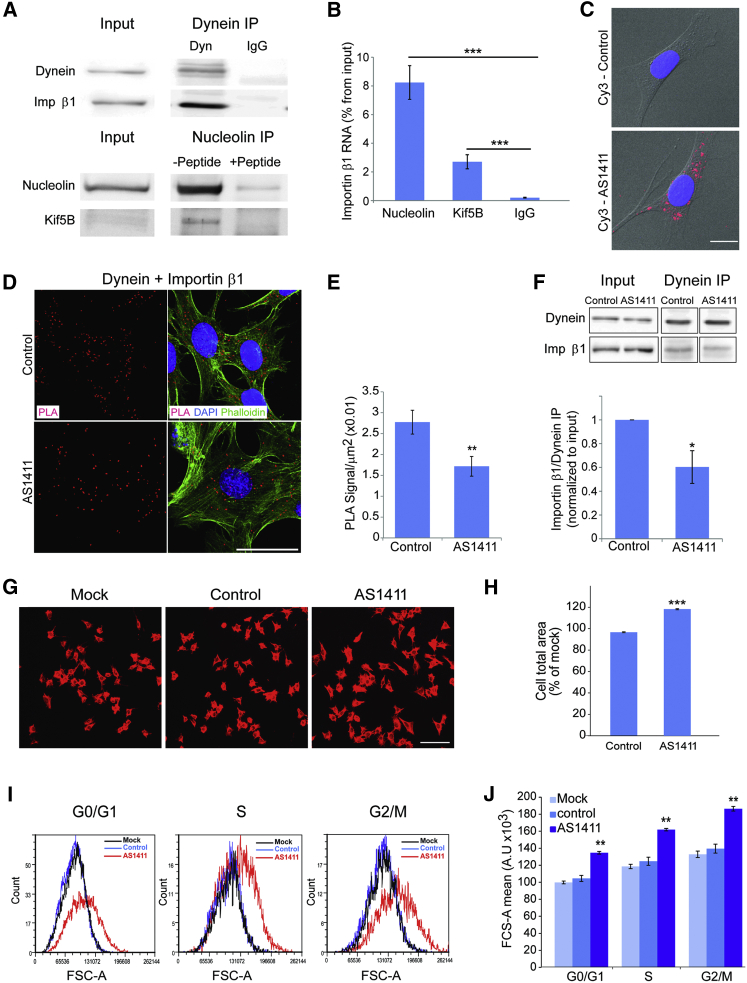
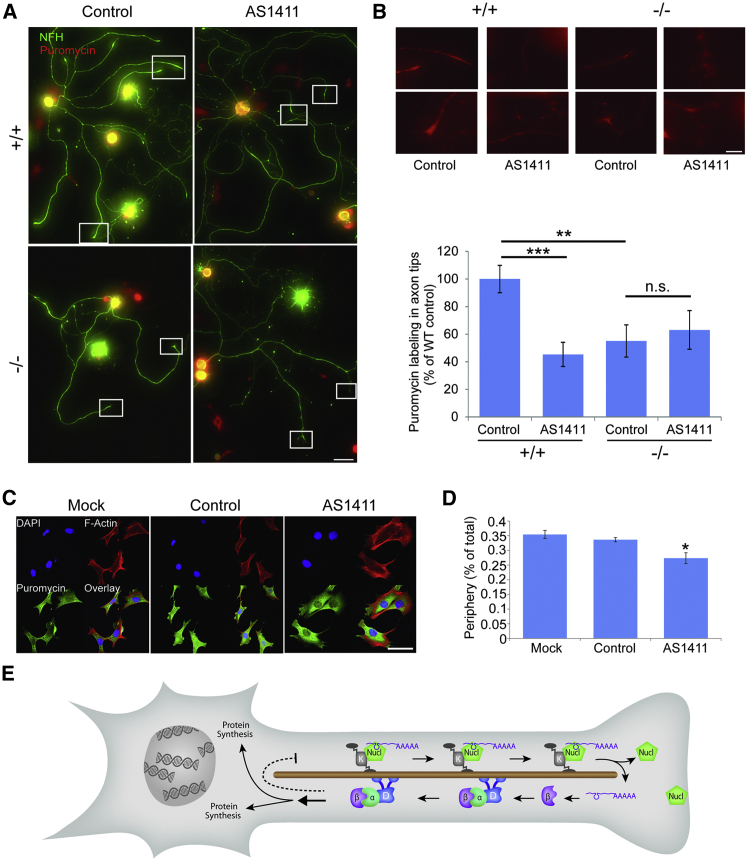
How can cells sense their own size to coordinate biosynthesis and metabolism with their growth needs? We recently proposed a motor-dependent bidirectional transport mechanism for axon length and cell size sensing, but the nature of the motor-transported size signals remained elusive. Here, we show that motor-dependent mRNA localization regulates neuronal growth and cycling cell size. We found that the RNA-binding protein nucleolin is associated with importin β1 mRNA in axons. Perturbation of nucleolin association with kinesins reduces its levels in axons, with a concomitant reduction in axonal importin β1 mRNA and protein levels. Strikingly, subcellular sequestration of nucleolin or importin β1 enhances axonal growth and causes a subcellular shift in protein synthesis. Similar findings were obtained in fibroblasts. Thus, subcellular mRNA localization regulates size and growth in both neurons and cycling cells.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
The glycine arginine-rich domain of the RNA-binding protein nucleolin regulates its subcellular localization.EMBO J. 2021 Oct 18;40(20):e107158. doi: 10.15252/embj.2020107158. Epub 2021 Sep 13. EMBO J. 2021. PMID: 34515347 Free PMC article.
-
Locally translated mTOR controls axonal local translation in nerve injury.Science. 2018 Mar 23;359(6382):1416-1421. doi: 10.1126/science.aan1053. Science. 2018. PMID: 29567716 Free PMC article.
-
Dynamics of survival of motor neuron (SMN) protein interaction with the mRNA-binding protein IMP1 facilitates its trafficking into motor neuron axons.Dev Neurobiol. 2014 Mar;74(3):319-332. doi: 10.1002/dneu.22111. Epub 2013 Oct 4. Dev Neurobiol. 2014. PMID: 23897586 Free PMC article.
-
Molecular mechanisms behind mRNA localization in axons.Open Biol. 2020 Sep;10(9):200177. doi: 10.1098/rsob.200177. Epub 2020 Sep 23. Open Biol. 2020. PMID: 32961072 Free PMC article. Review.
-
Molecular control of local translation in axon development and maintenance.Curr Opin Neurobiol. 2018 Aug;51:86-94. doi: 10.1016/j.conb.2018.02.025. Epub 2018 Mar 14. Curr Opin Neurobiol. 2018. PMID: 29549711 Review.
Cited by
-
A conditional null allele of Dync1h1 enables targeted analyses of dynein roles in neuronal length sensing.J Cell Sci. 2023 Mar 1;136(5):jcs260220. doi: 10.1242/jcs.260220. Epub 2022 Oct 31. J Cell Sci. 2023. PMID: 36218033 Free PMC article.
-
The Development of Mechanical Allodynia in Diabetic Rats Revealed by Single-Cell RNA-Seq.Front Mol Neurosci. 2022 May 20;15:856299. doi: 10.3389/fnmol.2022.856299. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35668789 Free PMC article.
-
A Ca2+-Dependent Switch Activates Axonal Casein Kinase 2α Translation and Drives G3BP1 Granule Disassembly for Axon Regeneration.Curr Biol. 2020 Dec 21;30(24):4882-4895.e6. doi: 10.1016/j.cub.2020.09.043. Epub 2020 Oct 15. Curr Biol. 2020. PMID: 33065005 Free PMC article.
-
Nucleolin: a cell portal for viruses, bacteria, and toxins.Cell Mol Life Sci. 2022 May 3;79(5):271. doi: 10.1007/s00018-022-04300-7. Cell Mol Life Sci. 2022. PMID: 35503380 Free PMC article. Review.
-
DYNLRB1 is essential for dynein mediated transport and neuronal survival.Neurobiol Dis. 2020 Jul;140:104816. doi: 10.1016/j.nbd.2020.104816. Epub 2020 Feb 20. Neurobiol Dis. 2020. PMID: 32088381 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
