

Opposing Functions of the N-terminal Acetyltransferases Naa50 and NatA in Sister-chromatid Cohesion
- PMID: 27422821
- PMCID: PMC5009278
- DOI: 10.1074/jbc.M116.737585
Opposing Functions of the N-terminal Acetyltransferases Naa50 and NatA in Sister-chromatid Cohesion
Abstract
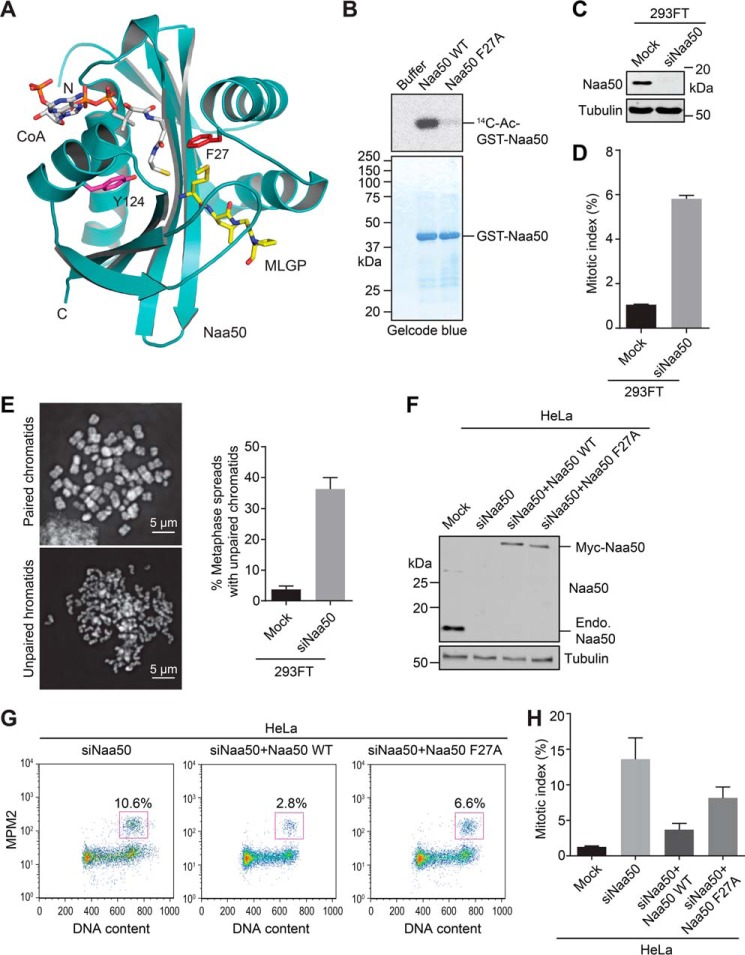
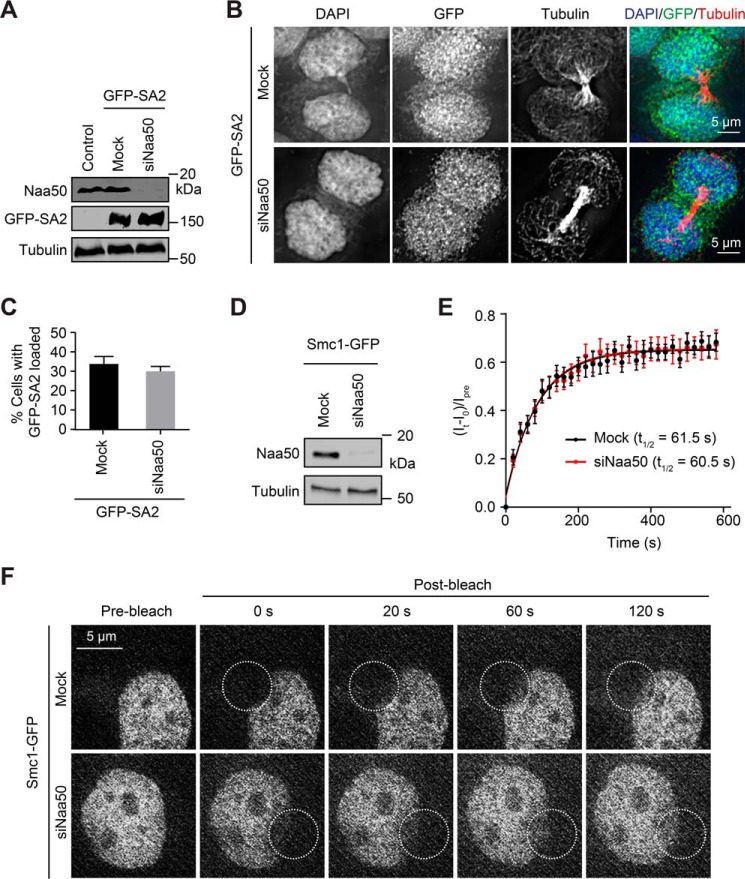
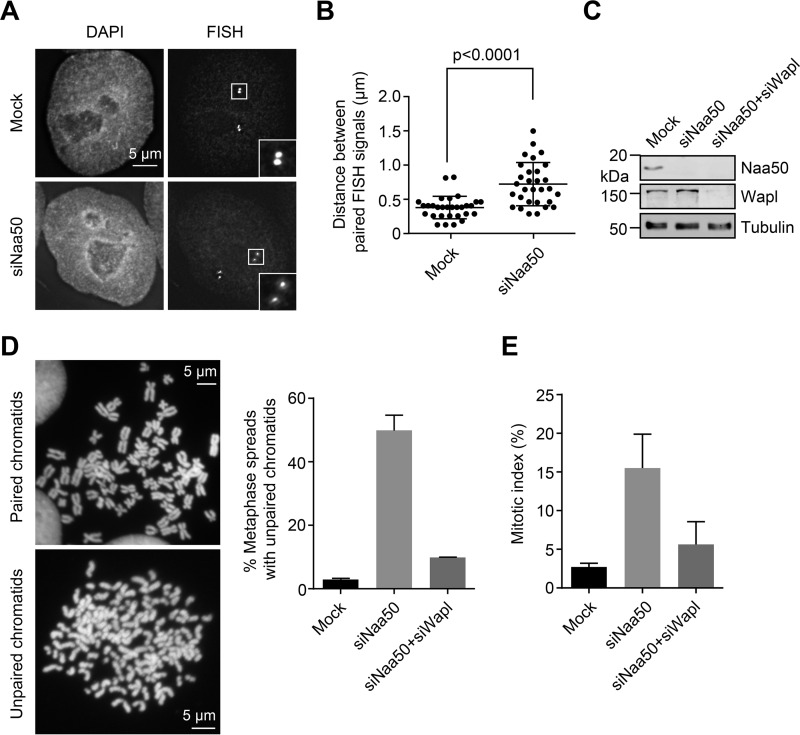
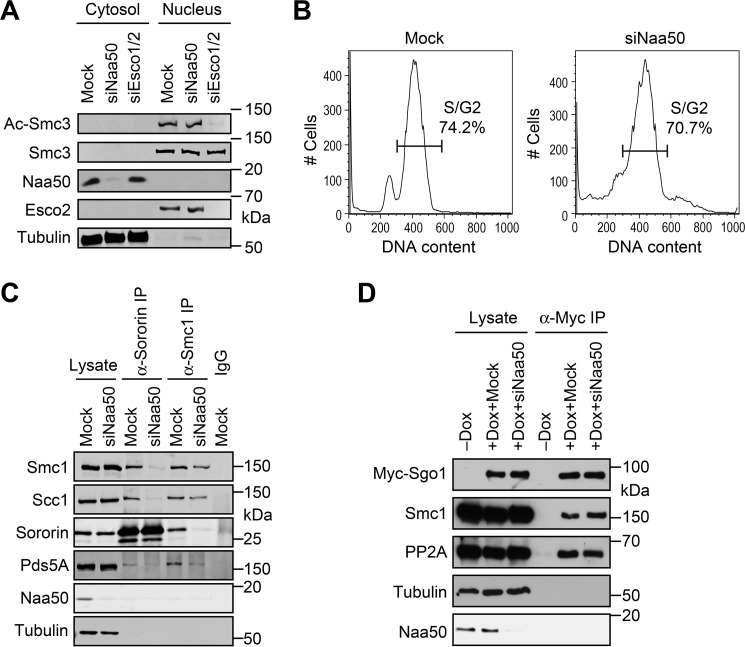
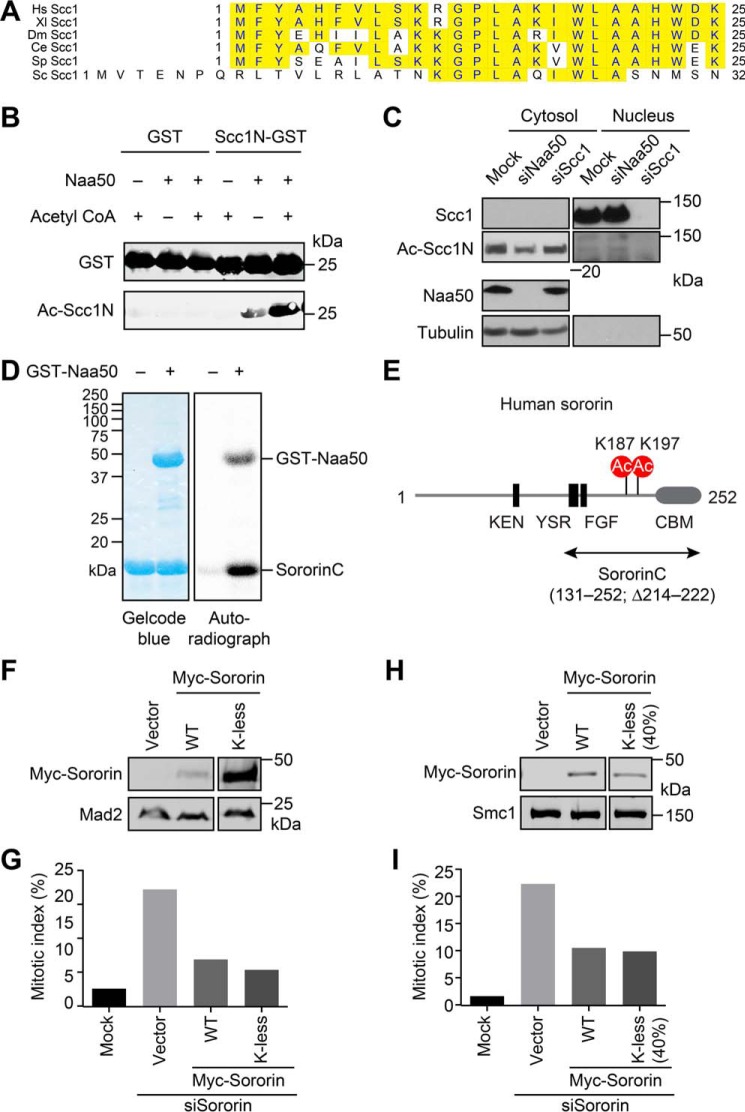
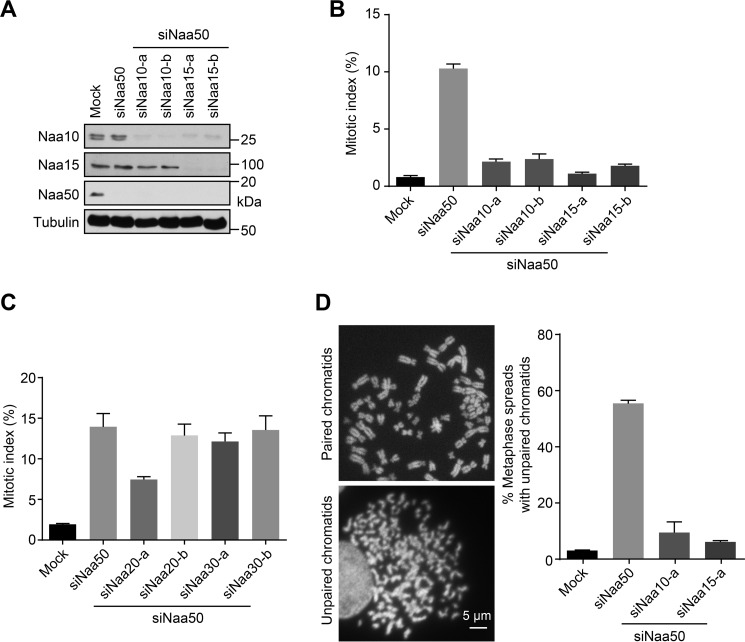
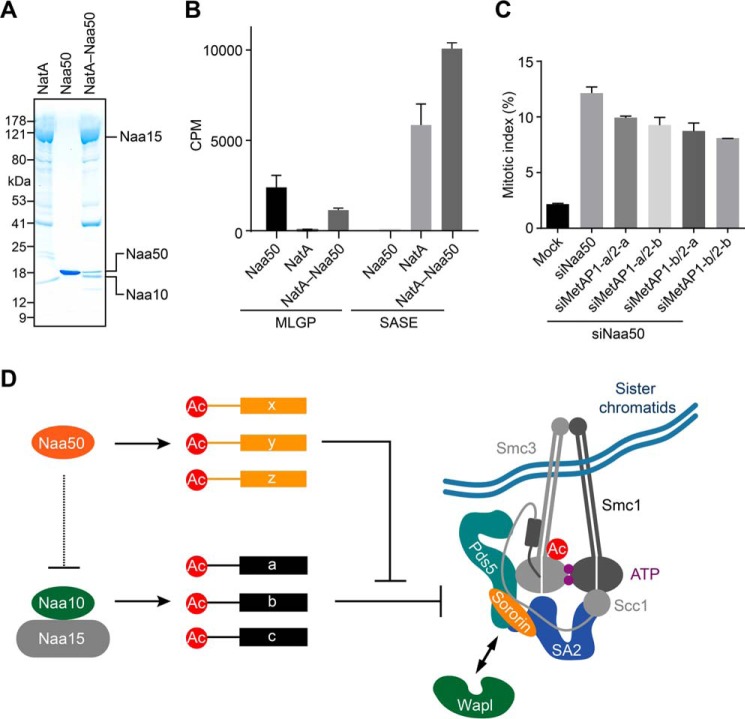
During the cell cycle, sister-chromatid cohesion tethers sister chromatids together from S phase to the metaphase-anaphase transition and ensures accurate segregation of chromatids into daughter cells. N-terminal acetylation is one of the most prevalent protein covalent modifications in eukaryotes and is mediated by a family of N-terminal acetyltransferases (NAT). Naa50 (also called San) has previously been shown to play a role in sister-chromatid cohesion in metazoans. The mechanism by which Naa50 contributes to cohesion is not understood however. Here, we show that depletion of Naa50 in HeLa cells weakens the interaction between cohesin and its positive regulator sororin and causes cohesion defects in S phase, consistent with a role of Naa50 in cohesion establishment. Strikingly, co-depletion of NatA, a heterodimeric NAT complex that physically interacts with Naa50, rescues the sister-chromatid cohesion defects and the resulting mitotic arrest caused by Naa50 depletion, indicating that NatA and Naa50 play antagonistic roles in cohesion. Purified recombinant NatA and Naa50 do not affect each other's NAT activity in vitro Because NatA and Naa50 exhibit distinct substrate specificity, we propose that they modify different effectors and regulate sister-chromatid cohesion in opposing ways.
Keywords: N-terminal acetylation; acetyltransferase; cell cycle; chromosomes; cohesin; enzyme; mitosis; sister-chromatid cohesion.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
The acetyltransferase activity of San stabilizes the mitotic cohesin at the centromeres in a shugoshin-independent manner.J Cell Biol. 2007 May 21;177(4):587-97. doi: 10.1083/jcb.200701043. Epub 2007 May 14. J Cell Biol. 2007. PMID: 17502424 Free PMC article.
-
Naa50/San-dependent N-terminal acetylation of Scc1 is potentially important for sister chromatid cohesion.Sci Rep. 2016 Dec 20;6:39118. doi: 10.1038/srep39118. Sci Rep. 2016. PMID: 27996020 Free PMC article.
-
The acetyltransferase Eco1 elicits cohesin dimerization during S phase.J Biol Chem. 2020 May 29;295(22):7554-7565. doi: 10.1074/jbc.RA120.013102. Epub 2020 Apr 20. J Biol Chem. 2020. PMID: 32312753 Free PMC article.
-
The expanding phenotypes of cohesinopathies: one ring to rule them all!Cell Cycle. 2019 Nov;18(21):2828-2848. doi: 10.1080/15384101.2019.1658476. Epub 2019 Sep 13. Cell Cycle. 2019. PMID: 31516082 Free PMC article. Review.
-
Cohesin complexes and sister chromatid cohesion in mammalian meiosis.Genome Dyn. 2009;5:94-116. doi: 10.1159/000166622. Genome Dyn. 2009. PMID: 18948710 Review.
Cited by
-
Structure of Human NatA and Its Regulation by the Huntingtin Interacting Protein HYPK.Structure. 2018 Jul 3;26(7):925-935.e8. doi: 10.1016/j.str.2018.04.003. Epub 2018 May 10. Structure. 2018. PMID: 29754825 Free PMC article.
-
A dual-function SNF2 protein drives chromatid resolution and nascent transcripts removal in mitosis.EMBO Rep. 2023 Sep 6;24(9):e56463. doi: 10.15252/embr.202256463. Epub 2023 Jul 18. EMBO Rep. 2023. PMID: 37462213 Free PMC article.
-
Absence of the Spindle Assembly Checkpoint Restores Mitotic Fidelity upon Loss of Sister Chromatid Cohesion.Curr Biol. 2018 Sep 10;28(17):2837-2844.e3. doi: 10.1016/j.cub.2018.06.062. Epub 2018 Aug 16. Curr Biol. 2018. PMID: 30122528 Free PMC article.
-
Spotlight on protein N-terminal acetylation.Exp Mol Med. 2018 Jul 27;50(7):1-13. doi: 10.1038/s12276-018-0116-z. Exp Mol Med. 2018. PMID: 30054468 Free PMC article. Review.
-
Structure and Mechanism of Acetylation by the N-Terminal Dual Enzyme NatA/Naa50 Complex.Structure. 2019 Jul 2;27(7):1057-1070.e4. doi: 10.1016/j.str.2019.04.014. Epub 2019 May 30. Structure. 2019. PMID: 31155310 Free PMC article.
References
-
- Nasmyth K. (2002) Segregating sister genomes: the molecular biology of chromosome separation. Science 297, 559–565 - PubMed
-
- Onn I., Heidinger-Pauli J. M., Guacci V., Unal E., and Koshland D. E. (2008) Sister chromatid cohesion: a simple concept with a complex reality. Annu. Rev. Cell Dev. Biol. 24, 105–129 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
