

A comprehensive transcriptional map of primate brain development
- PMID: 27409810
- PMCID: PMC5325728
- DOI: 10.1038/nature18637
A comprehensive transcriptional map of primate brain development
Abstract
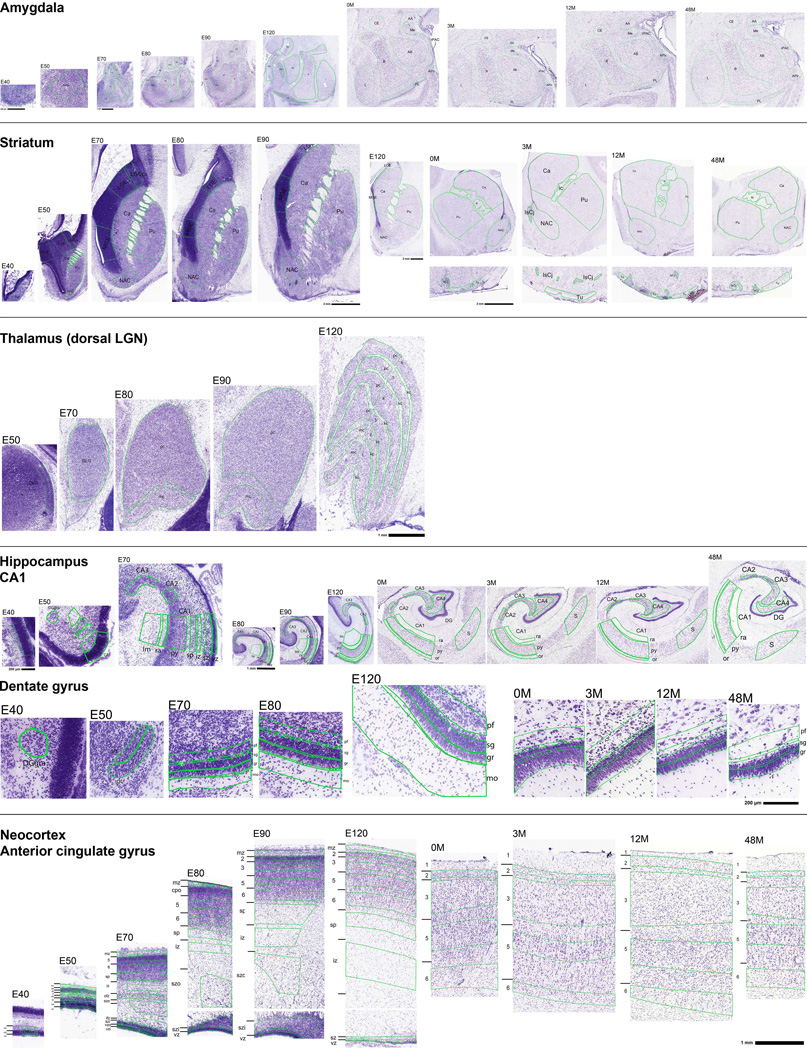
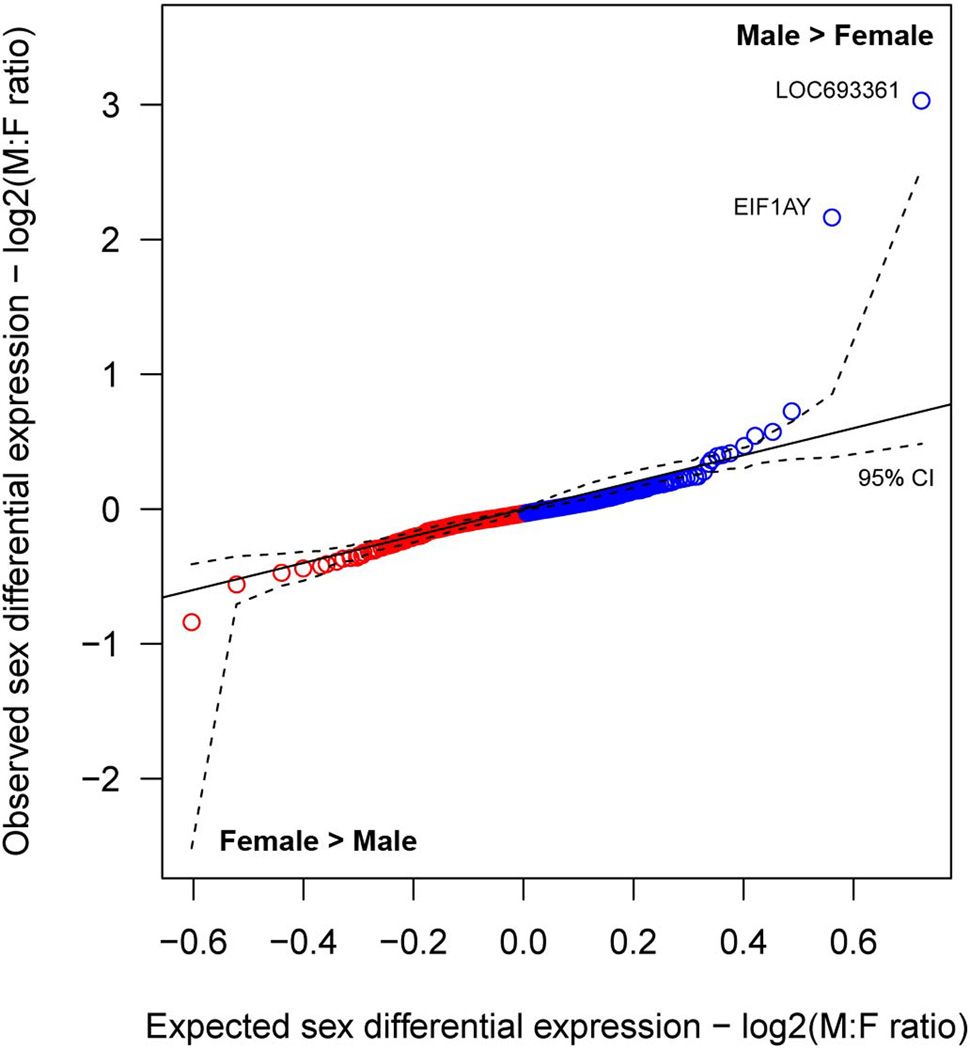
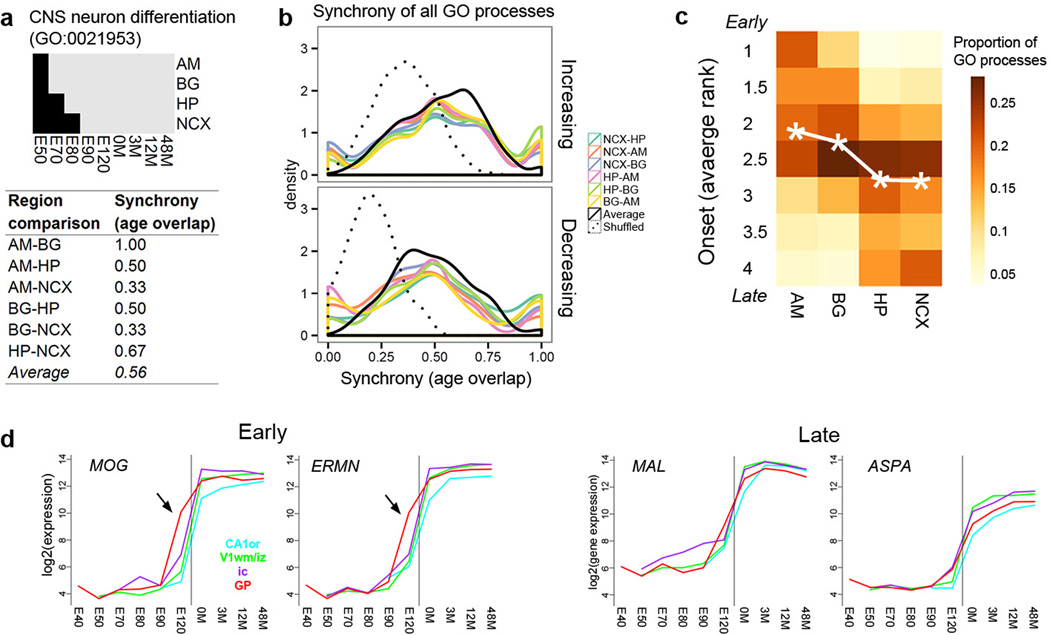
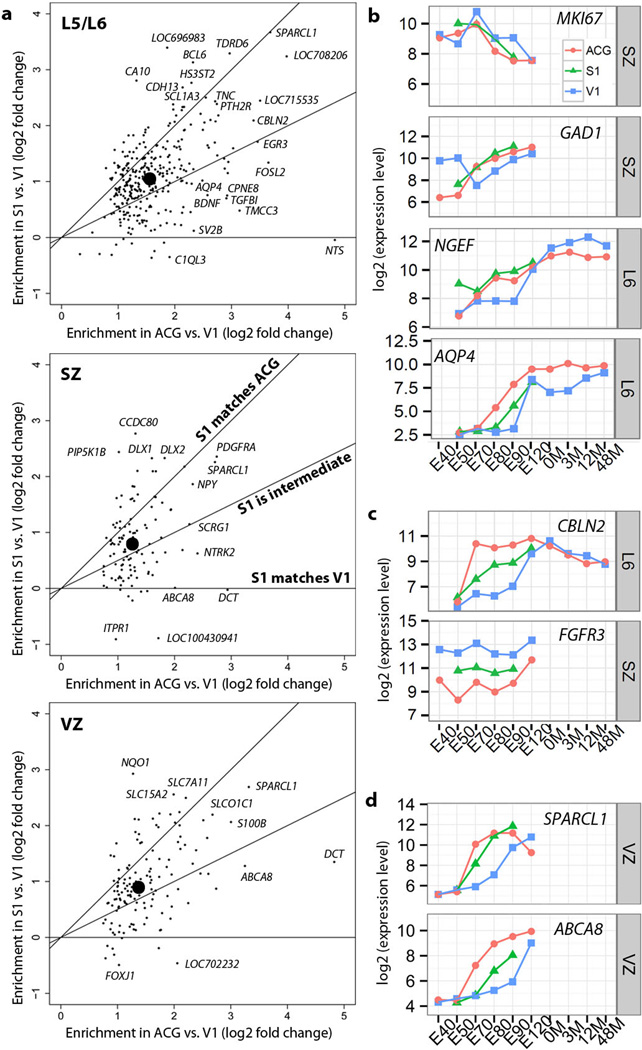
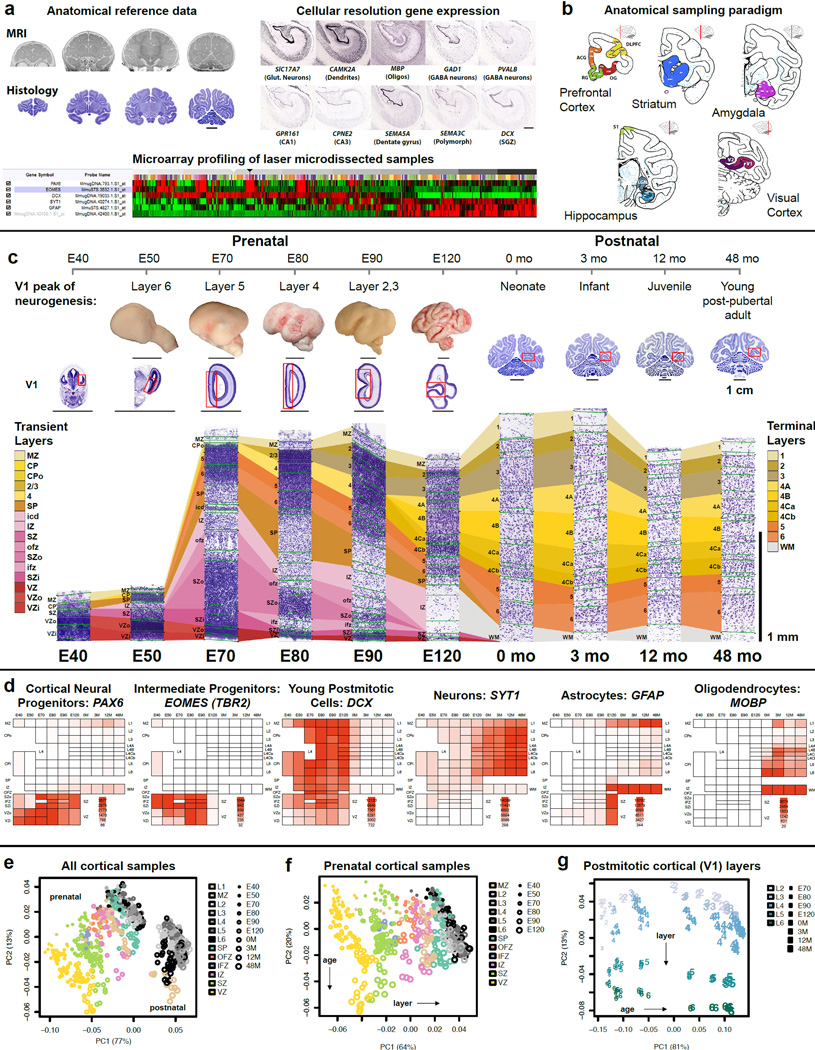
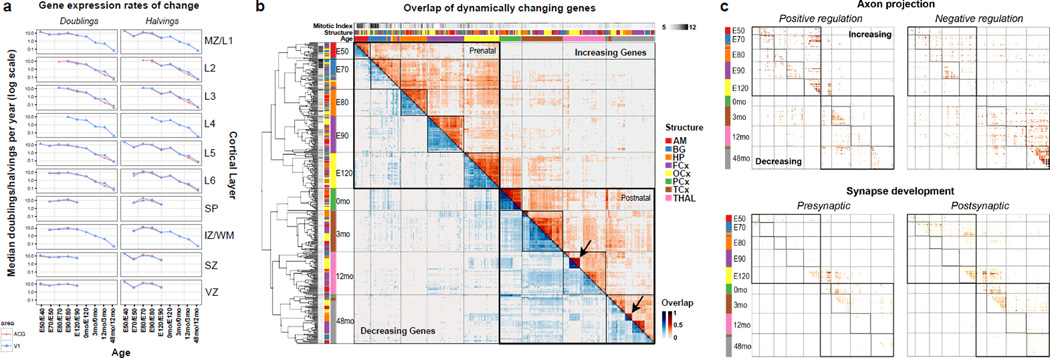
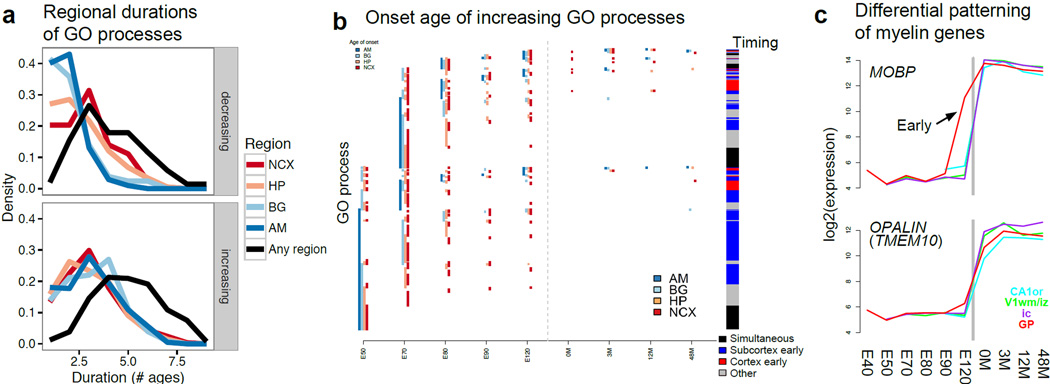
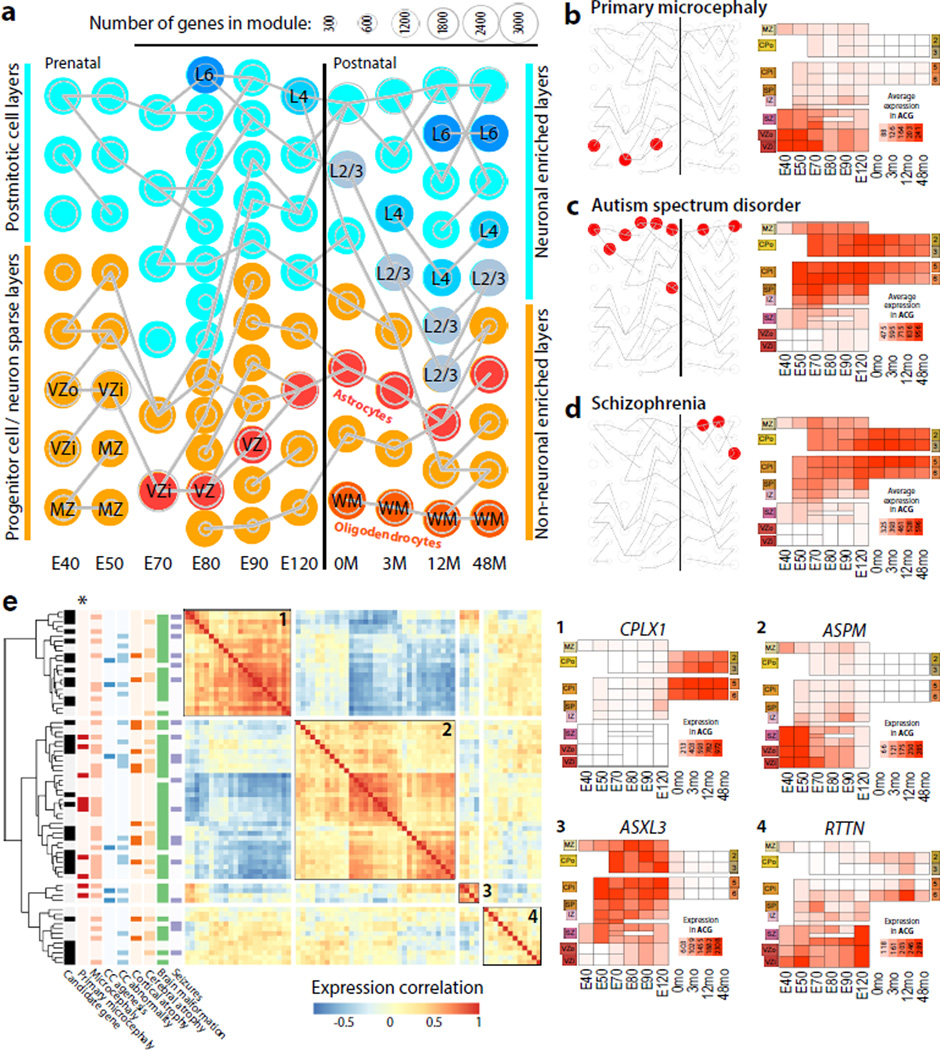
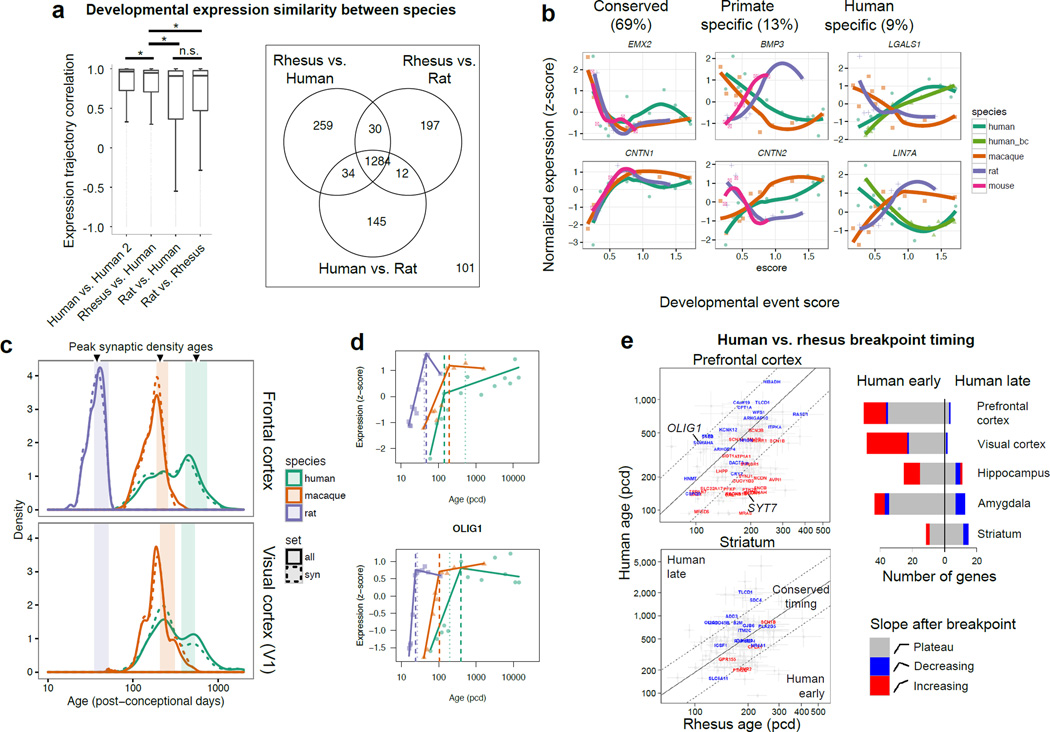
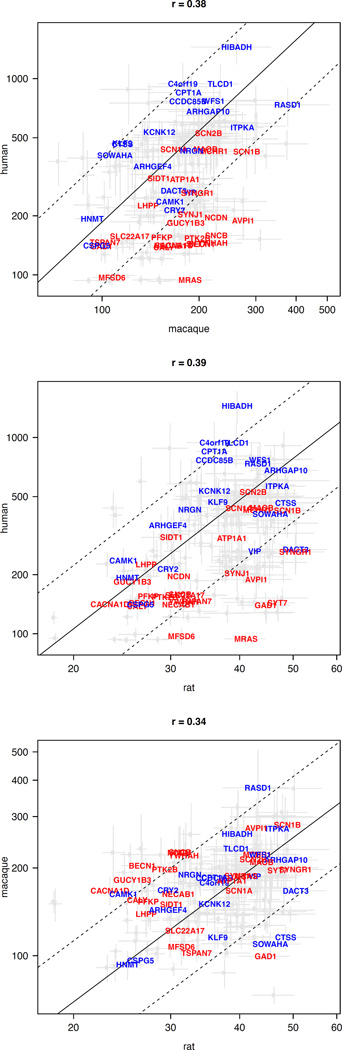
The transcriptional underpinnings of brain development remain poorly understood, particularly in humans and closely related non-human primates. We describe a high-resolution transcriptional atlas of rhesus monkey (Macaca mulatta) brain development that combines dense temporal sampling of prenatal and postnatal periods with fine anatomical division of cortical and subcortical regions associated with human neuropsychiatric disease. Gene expression changes more rapidly before birth, both in progenitor cells and maturing neurons. Cortical layers and areas acquire adult-like molecular profiles surprisingly late in postnatal development. Disparate cell populations exhibit distinct developmental timing of gene expression, but also unexpected synchrony of processes underlying neural circuit construction including cell projection and adhesion. Candidate risk genes for neurodevelopmental disorders including primary microcephaly, autism spectrum disorder, intellectual disability, and schizophrenia show disease-specific spatiotemporal enrichment within developing neocortex. Human developmental expression trajectories are more similar to monkey than rodent, although approximately 9% of genes show human-specific regulation with evidence for prolonged maturation or neoteny compared to monkey.
Figures



Similar articles
-
Spatiotemporal dynamics of the postnatal developing primate brain transcriptome.Hum Mol Genet. 2015 Aug 1;24(15):4327-39. doi: 10.1093/hmg/ddv166. Epub 2015 May 7. Hum Mol Genet. 2015. PMID: 25954031 Free PMC article.
-
Spatiotemporal transcriptomic divergence across human and macaque brain development.Science. 2018 Dec 14;362(6420):eaat8077. doi: 10.1126/science.aat8077. Epub 2018 Dec 13. Science. 2018. PMID: 30545855 Free PMC article.
-
Transcriptomic signatures of neuronal differentiation and their association with risk genes for autism spectrum and related neuropsychiatric disorders.Transl Psychiatry. 2016 Aug 2;6(8):e864. doi: 10.1038/tp.2016.119. Transl Psychiatry. 2016. PMID: 27483382 Free PMC article.
-
The Pleiotropic MET Receptor Network: Circuit Development and the Neural-Medical Interface of Autism.Biol Psychiatry. 2017 Mar 1;81(5):424-433. doi: 10.1016/j.biopsych.2016.08.035. Epub 2016 Sep 15. Biol Psychiatry. 2017. PMID: 27837921 Free PMC article. Review.
-
Transcriptional Regulators and Human-Specific/Primate-Specific Genes in Neocortical Neurogenesis.Int J Mol Sci. 2020 Jun 29;21(13):4614. doi: 10.3390/ijms21134614. Int J Mol Sci. 2020. PMID: 32610533 Free PMC article. Review.
Cited by
-
Non-Human Primate Blood-Brain Barrier and In Vitro Brain Endothelium: From Transcriptome to the Establishment of a New Model.Pharmaceutics. 2020 Oct 14;12(10):967. doi: 10.3390/pharmaceutics12100967. Pharmaceutics. 2020. PMID: 33066641 Free PMC article.
-
What Makes Organoids Good Models of Human Neurogenesis?Front Neurosci. 2022 Apr 14;16:872794. doi: 10.3389/fnins.2022.872794. eCollection 2022. Front Neurosci. 2022. PMID: 35495031 Free PMC article.
-
Multimodal Single-Cell Analysis Reveals Physiological Maturation in the Developing Human Neocortex.Neuron. 2019 Apr 3;102(1):143-158.e7. doi: 10.1016/j.neuron.2019.01.027. Epub 2019 Feb 12. Neuron. 2019. PMID: 30770253 Free PMC article.
-
Development and disease in a dish: the epigenetics of neurodevelopmental disorders.Epigenomics. 2018 Feb;10(2):219-231. doi: 10.2217/epi-2017-0113. Epub 2018 Jan 15. Epigenomics. 2018. PMID: 29334242 Free PMC article. Review.
-
Profiling gene expression in the human dentate gyrus granule cell layer reveals insights into schizophrenia and its genetic risk.Nat Neurosci. 2020 Apr;23(4):510-519. doi: 10.1038/s41593-020-0604-z. Epub 2020 Mar 16. Nat Neurosci. 2020. PMID: 32203495
References
-
- Caldwell CA, Whiten Ain. Primates Perspect. Oxford University Press; 2011. pp. 652–662.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
