

RBPJ Controls Development of Pathogenic Th17 Cells by Regulating IL-23 Receptor Expression
- PMID: 27346359
- PMCID: PMC4984261
- DOI: 10.1016/j.celrep.2016.05.088
RBPJ Controls Development of Pathogenic Th17 Cells by Regulating IL-23 Receptor Expression
Abstract
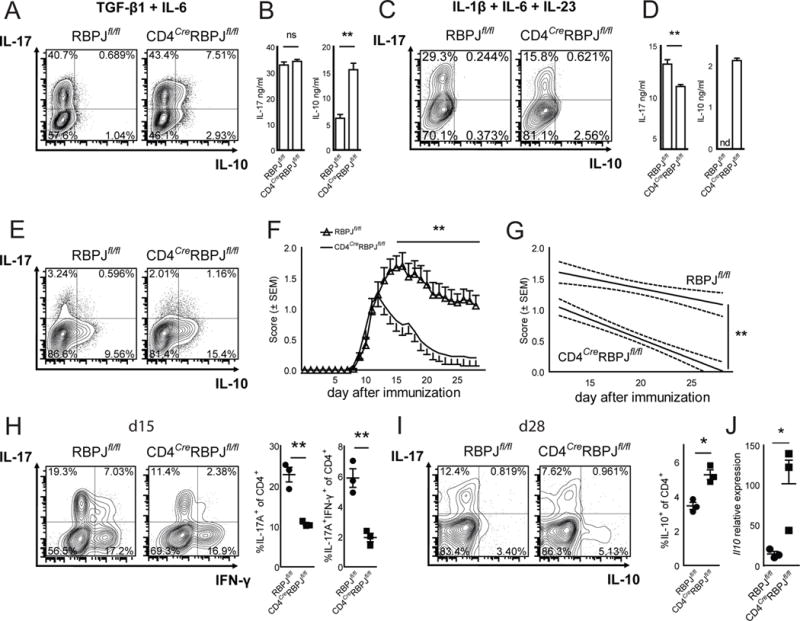
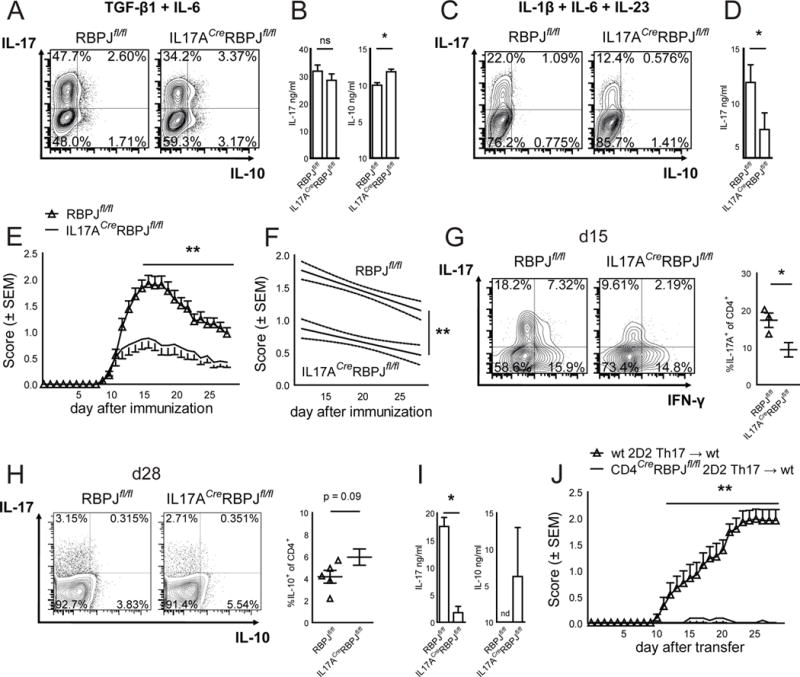
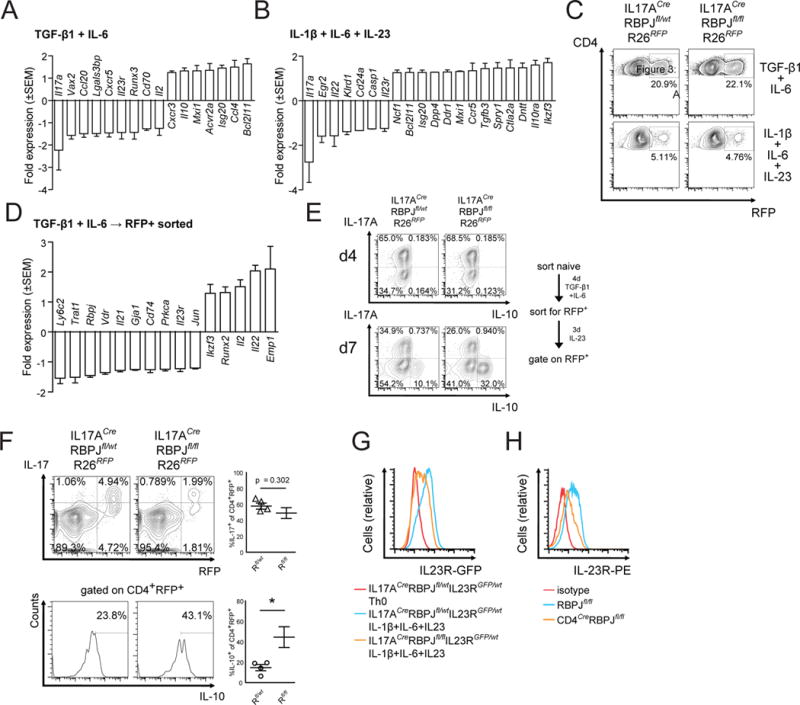
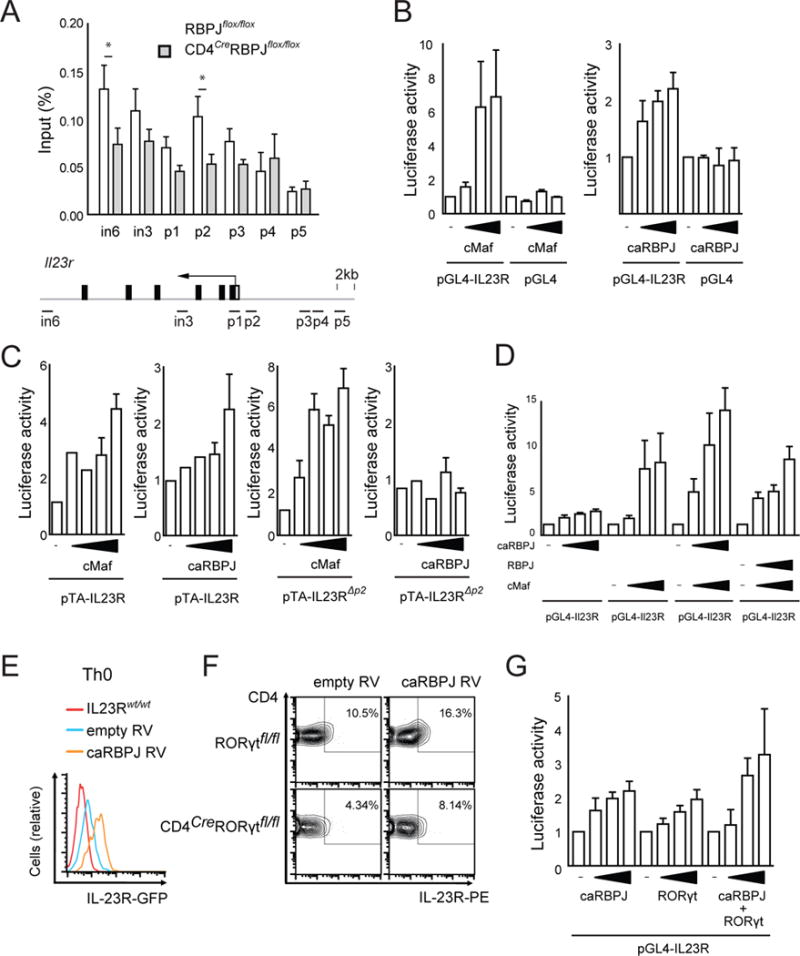
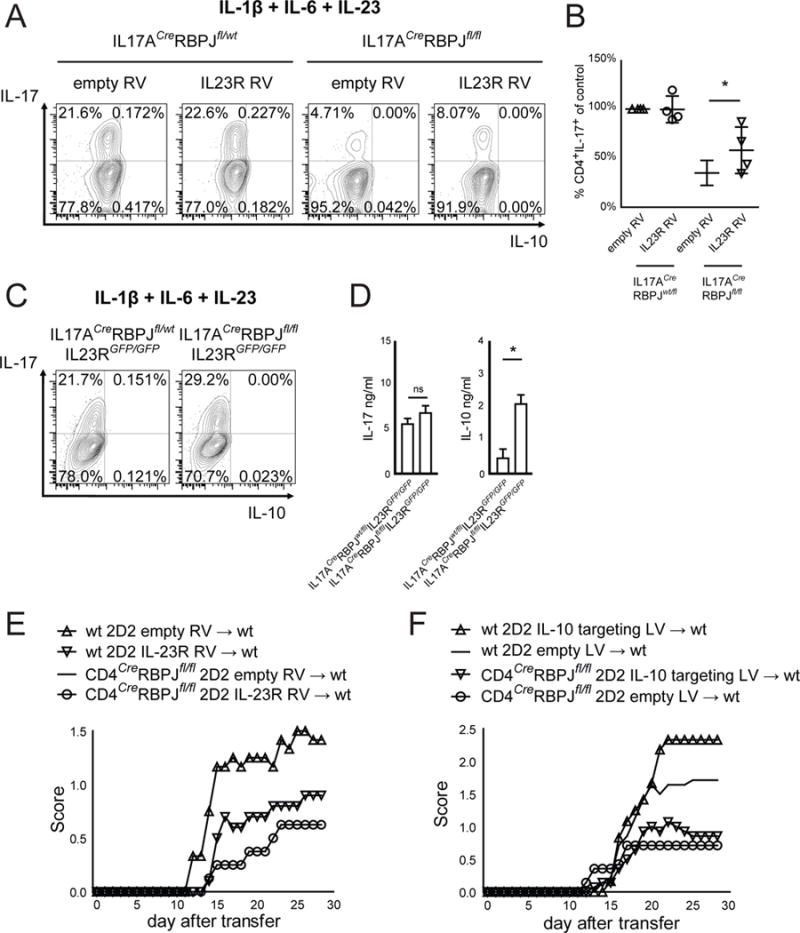
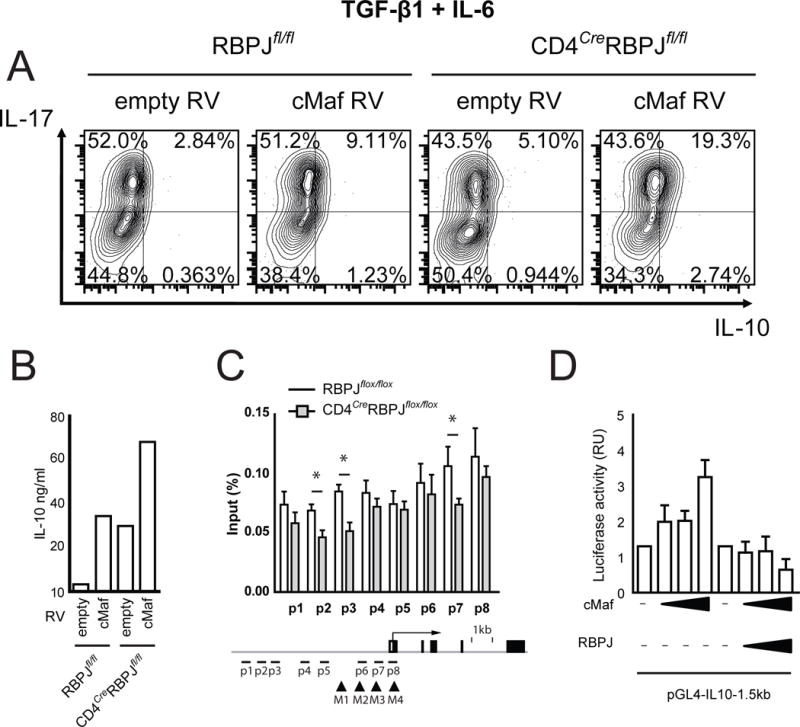
Interleukin-17 (IL-17)-producing helper T cells (Th17 cells) play an important role in autoimmune diseases. However, not all Th17 cells induce tissue inflammation or autoimmunity. Th17 cells require IL-23 receptor (IL-23R) signaling to become pathogenic. The transcriptional mechanisms controlling the pathogenicity of Th17 cells and IL-23R expression are unknown. Here, we demonstrate that the canonical Notch signaling mediator RBPJ is a key driver of IL-23R expression. In the absence of RBPJ, Th17 cells fail to upregulate IL-23R, lack stability, and do not induce autoimmune tissue inflammation in vivo, whereas overexpression of IL-23R rescues this defect and promotes pathogenicity of RBPJ-deficient Th17 cells. RBPJ binds and trans-activates the Il23r promoter and induces IL-23R expression and represses anti-inflammatory IL-10 production in Th17 cells. We thus find that Notch signaling influences the development of pathogenic and non-pathogenic Th17 cells by reciprocally regulating IL-23R and IL-10 expression.
Keywords: IL-23R; Notch; RBPJ; Th17 cells; pathogenicity.
Published by Elsevier Inc.
Figures
Similar articles
-
IL-7/IL-7 Receptor Signaling Differentially Affects Effector CD4+ T Cell Subsets Involved in Experimental Autoimmune Encephalomyelitis.J Immunol. 2015 Sep 1;195(5):1974-83. doi: 10.4049/jimmunol.1403135. Epub 2015 Jul 29. J Immunol. 2015. PMID: 26223651 Free PMC article.
-
Th17 cell promotes apoptosis of IL-23R+ neurons in experimental autoimmune encephalomyelitis.Clin Immunol. 2024 Feb;259:109898. doi: 10.1016/j.clim.2024.109898. Epub 2024 Jan 5. Clin Immunol. 2024. PMID: 38185267
-
Nuclear receptor NR4A2 orchestrates Th17 cell-mediated autoimmune inflammation via IL-21 signalling.PLoS One. 2013;8(2):e56595. doi: 10.1371/journal.pone.0056595. Epub 2013 Feb 21. PLoS One. 2013. PMID: 23437182 Free PMC article.
-
Protective role of R381Q (rs11209026) polymorphism in IL-23R gene in immune-mediated diseases: A comprehensive review.J Immunotoxicol. 2016 May;13(3):286-300. doi: 10.3109/1547691X.2015.1115448. Epub 2016 Apr 4. J Immunotoxicol. 2016. PMID: 27043356 Review.
-
Molecular control of pathogenic Th17 cells in autoimmune diseases.Int Immunopharmacol. 2020 Mar;80:106187. doi: 10.1016/j.intimp.2020.106187. Epub 2020 Jan 10. Int Immunopharmacol. 2020. PMID: 31931372 Free PMC article. Review.
Cited by
-
The role of the Notch signalling pathway in the pathogenesis of ulcerative colitis: from the perspective of intestinal mucosal barrier.Front Med (Lausanne). 2024 Jan 5;10:1333531. doi: 10.3389/fmed.2023.1333531. eCollection 2023. Front Med (Lausanne). 2024. PMID: 38249980 Free PMC article. Review.
-
Insight Into Non-Pathogenic Th17 Cells in Autoimmune Diseases.Front Immunol. 2018 May 28;9:1112. doi: 10.3389/fimmu.2018.01112. eCollection 2018. Front Immunol. 2018. PMID: 29892286 Free PMC article. Review.
-
Emerging role of C5a/C5aR IL-17A axis in cGVHD.Am J Transl Res. 2018 Jul 15;10(7):2148-2157. eCollection 2018. Am J Transl Res. 2018. PMID: 30093951 Free PMC article.
-
Epigenetic and transcriptional mechanisms for the regulation of IL-10.Semin Immunol. 2019 Aug;44:101324. doi: 10.1016/j.smim.2019.101324. Epub 2019 Oct 30. Semin Immunol. 2019. PMID: 31676122 Free PMC article. Review.
-
Quantitative Proteomics Reveals the Dynamic Protein Landscape during Initiation of Human Th17 Cell Polarization.iScience. 2019 Jan 25;11:334-355. doi: 10.1016/j.isci.2018.12.020. Epub 2018 Dec 26. iScience. 2019. PMID: 30641411 Free PMC article.
References
-
- Amsen D, Antov A, Flavell RA. The different faces of Notch in T-helper-cell differentiation. Nature reviews Immunology. 2009;9:116–124. - PubMed
-
- Awasthi A, Carrier Y, Peron JP, Bettelli E, Kamanaka M, Flavell RA, Kuchroo VK, Oukka M, Weiner HL. A dominant function for interleukin 27 in generating interleukin 10-producing anti-inflammatory T cells. Nature immunology. 2007;8:1380–1389. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
