

Persistent Activation of NF-κB in BRCA1-Deficient Mammary Progenitors Drives Aberrant Proliferation and Accumulation of DNA Damage
- PMID: 27292187
- PMCID: PMC6850408
- DOI: 10.1016/j.stem.2016.05.003
Persistent Activation of NF-κB in BRCA1-Deficient Mammary Progenitors Drives Aberrant Proliferation and Accumulation of DNA Damage
Abstract
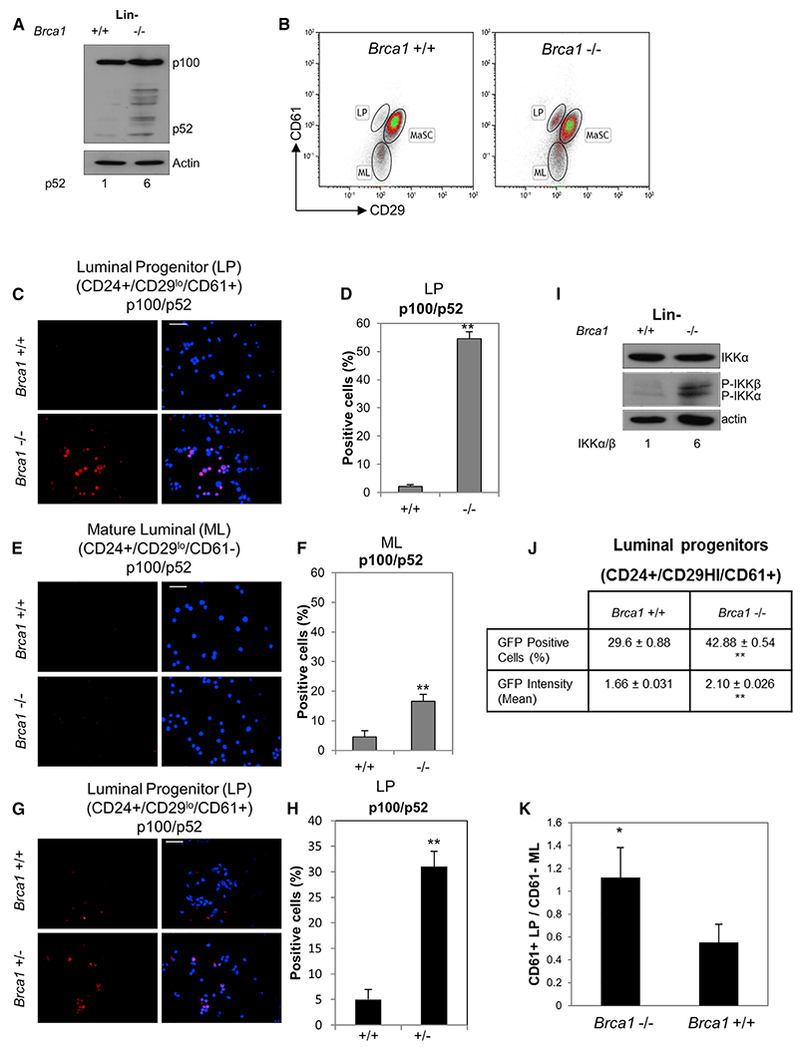
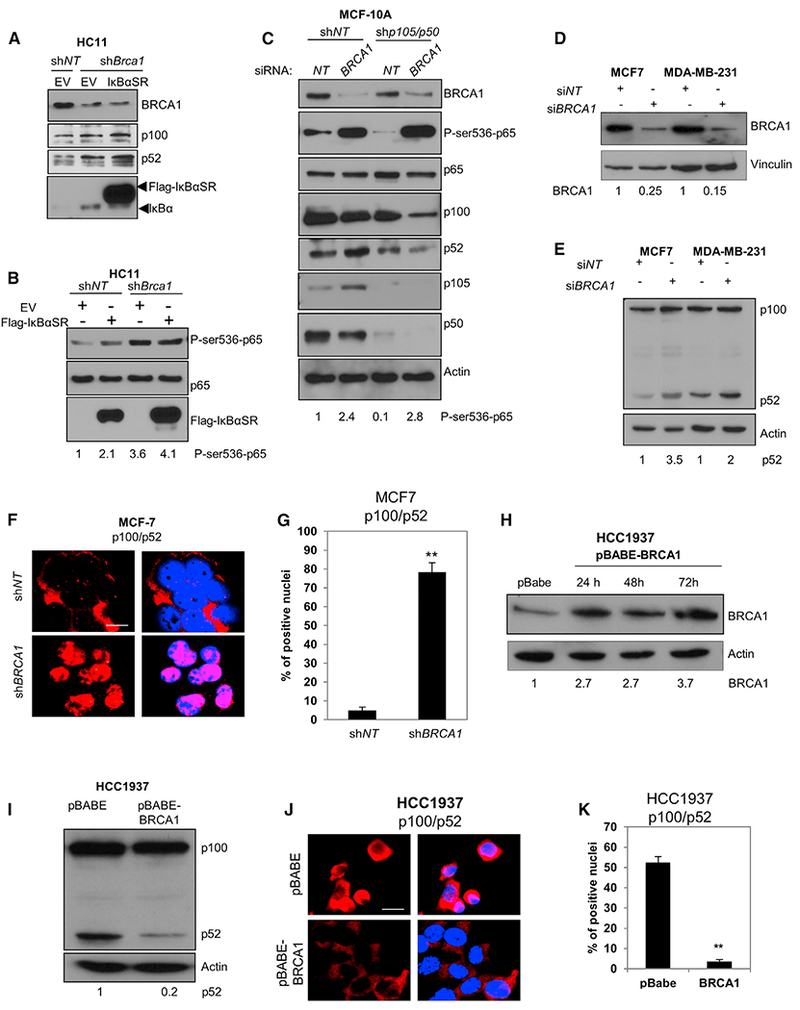
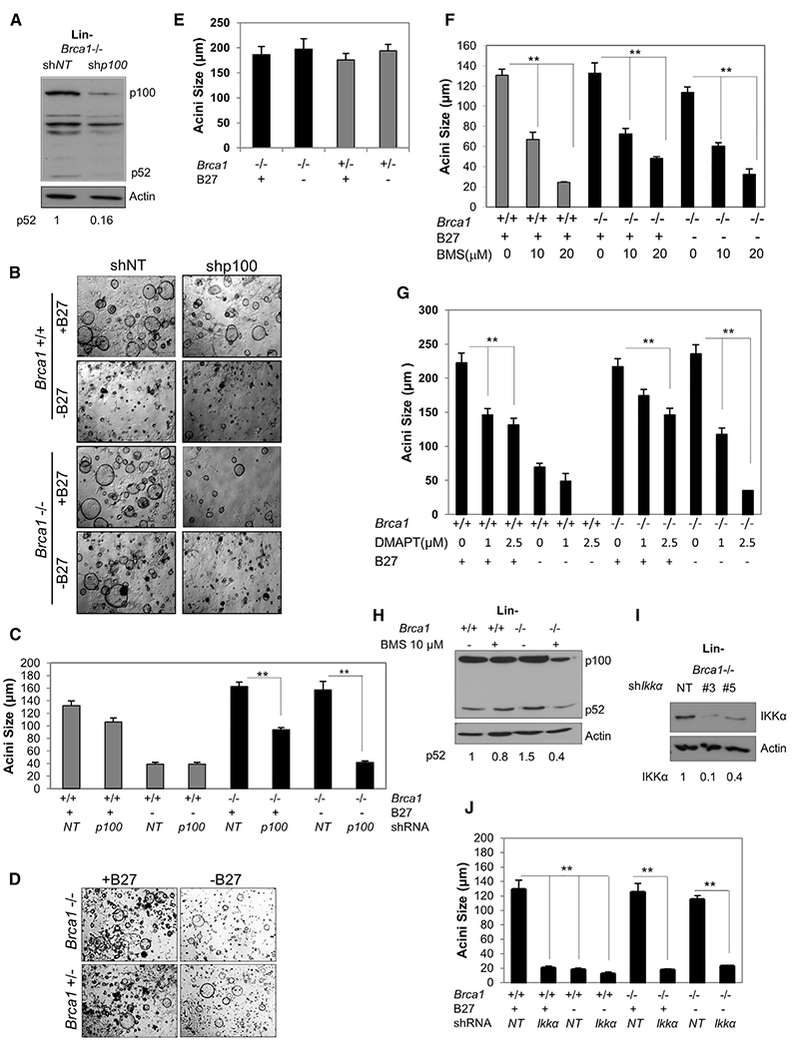
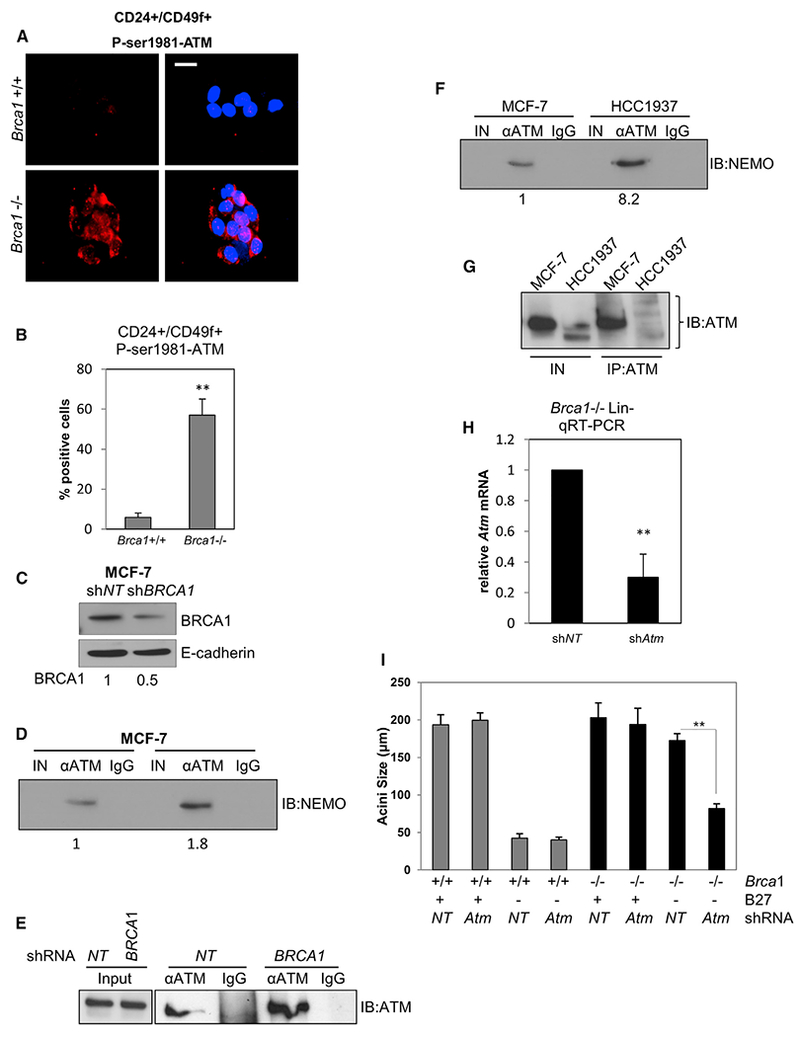
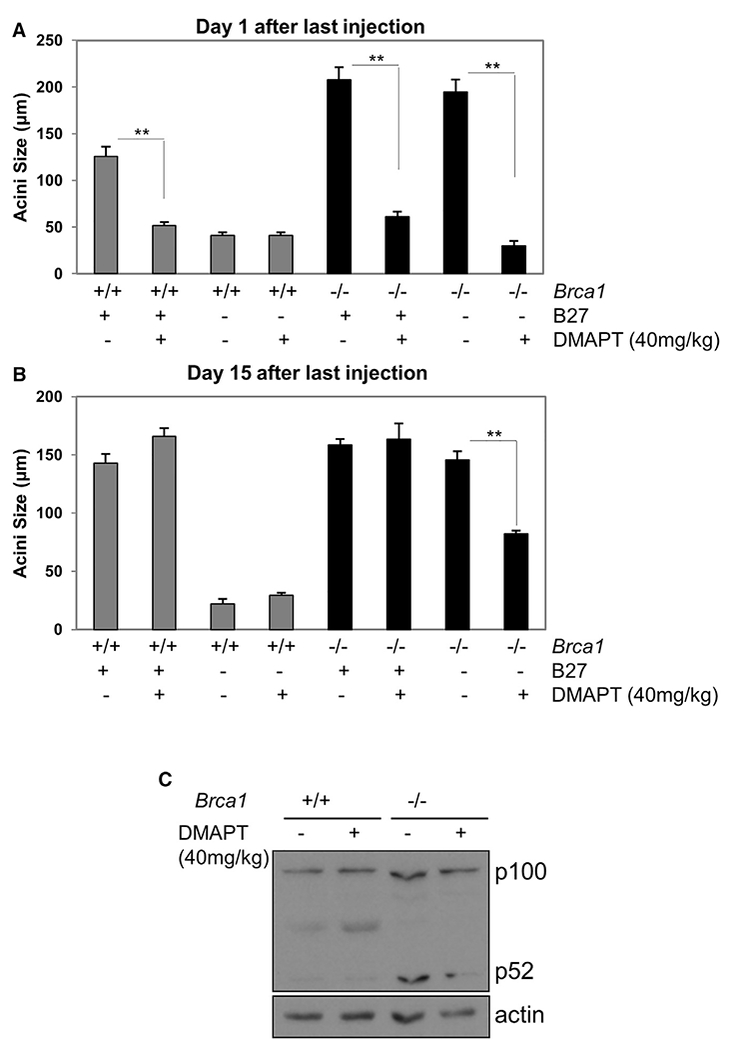
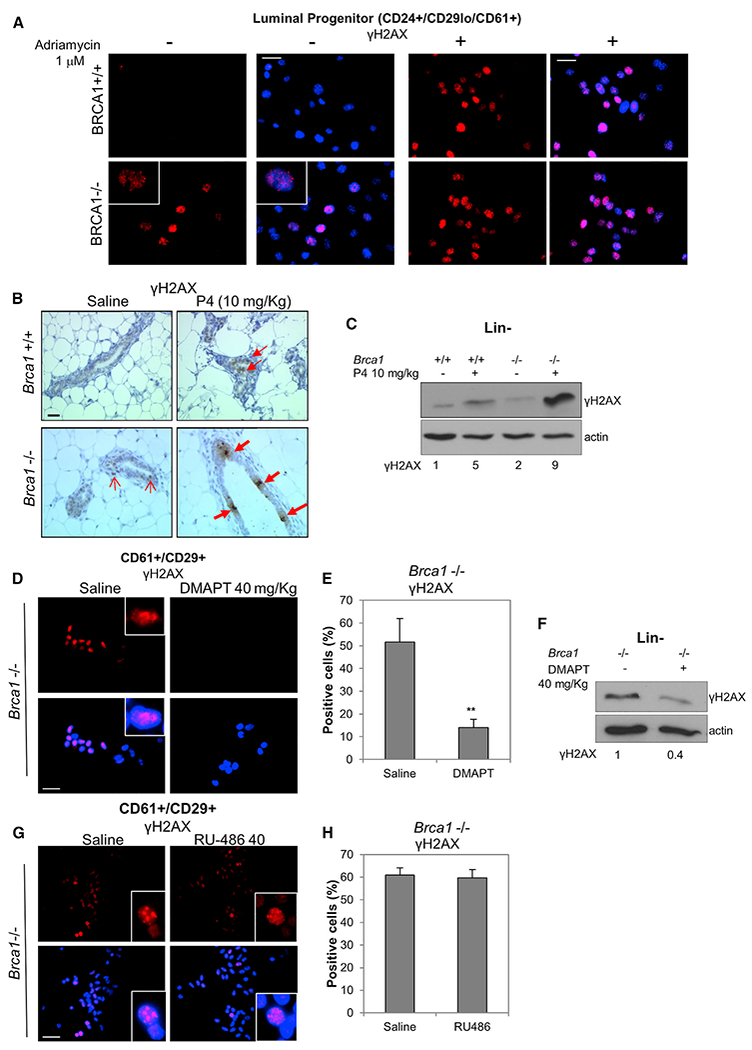
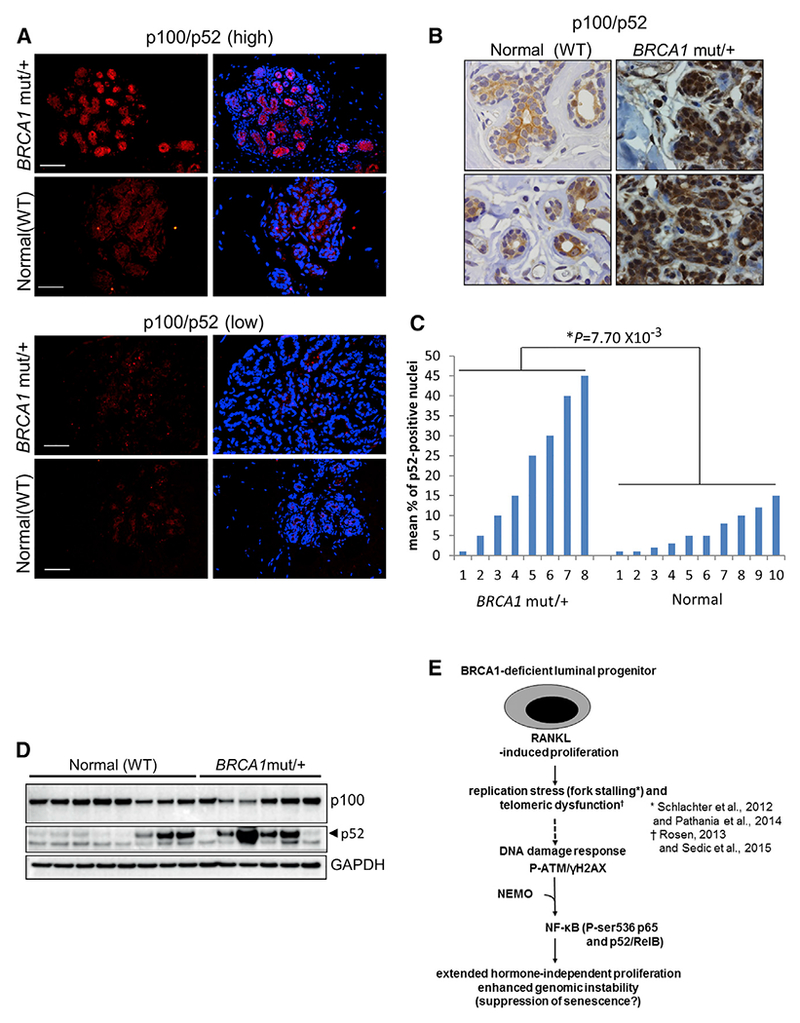
Human BRCA1 mutation carriers and BRCA1-deficient mouse mammary glands contain an abnormal population of mammary luminal progenitors that can form 3D colonies in a hormone-independent manner. The intrinsic cellular regulatory defect in these presumptive breast cancer precursors is not known. We have discovered that nuclear factor kappaB (NF-κB) (p52/RelB) is persistently activated in a subset of BRCA1-deficient mammary luminal progenitors. Hormone-independent luminal progenitor colony formation required NF-κB, ataxia telangiectasia-mutated (ATM), and the inhibitor of kappaB kinase, IKKα. Progesterone (P4)-stimulated proliferation resulted in a marked enhancement of DNA damage foci in Brca1(-/-) mouse mammary. In vivo, NF-κB inhibition prevented recovery of Brca1(-/-) hormone-independent colony-forming cells. The majority of human BRCA1(mut/+) mammary glands showed marked lobular expression of nuclear NF-κB. We conclude that the aberrant proliferative capacity of Brca1(-/-) luminal progenitor cells is linked to the replication-associated DNA damage response, where proliferation of mammary progenitors is perpetuated by damage-induced, autologous NF-κB signaling.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
CONFLICTS OF INTEREST
The University of Kentucky holds a patent on DMAPT. A potential royalty to P.A.C. may occur consistent with University of Kentucky policy. P.A.C. is a founder of Leuchemix, Inc.
Figures
Similar articles
-
NF-κB at the Crossroads of Normal Mammary Gland Biology and the Pathogenesis and Prevention of BRCA1-Mutated Breast Cancer.Cancer Prev Res (Phila). 2018 Feb;11(2):69-80. doi: 10.1158/1940-6207.CAPR-17-0225. Epub 2017 Nov 3. Cancer Prev Res (Phila). 2018. PMID: 29101208 Review.
-
RelB/p52 NF-kappaB complexes rescue an early delay in mammary gland development in transgenic mice with targeted superrepressor IkappaB-alpha expression and promote carcinogenesis of the mammary gland.Mol Cell Biol. 2005 Nov;25(22):10136-47. doi: 10.1128/MCB.25.22.10136-10147.2005. Mol Cell Biol. 2005. PMID: 16260626 Free PMC article.
-
c-Kit is required for growth and survival of the cells of origin of Brca1-mutation-associated breast cancer.Oncogene. 2012 Feb 16;31(7):869-83. doi: 10.1038/onc.2011.289. Epub 2011 Jul 18. Oncogene. 2012. PMID: 21765473
-
BRCA1 Attenuates Progesterone Effects on Proliferation and NFκB Activation in Normal Human Mammary Epithelial Cells.J Mammary Gland Biol Neoplasia. 2019 Sep;24(3):257-270. doi: 10.1007/s10911-019-09431-5. Epub 2019 May 18. J Mammary Gland Biol Neoplasia. 2019. PMID: 31104199
-
NF-kappaB in mammary gland development and breast cancer.J Mammary Gland Biol Neoplasia. 2003 Apr;8(2):215-23. doi: 10.1023/a:1025905008934. J Mammary Gland Biol Neoplasia. 2003. PMID: 14635796 Review.
Cited by
-
Associations of TRAF2 (rs867186), TAB2 (rs237025), IKBKB (rs13278372) Polymorphisms and TRAF2, TAB2, IKBKB Protein Levels with Clinical and Morphological Features of Pituitary Adenomas.Cancers (Basel). 2024 Jul 10;16(14):2509. doi: 10.3390/cancers16142509. Cancers (Basel). 2024. PMID: 39061149 Free PMC article.
-
Simultaneous Inhibition of EGFR and HER2 via Afatinib Augments the Radiosensitivity of Nasopharyngeal Carcinoma Cells.J Cancer. 2019 May 12;10(9):2063-2073. doi: 10.7150/jca.29327. eCollection 2019. J Cancer. 2019. PMID: 31205567 Free PMC article.
-
Stem Cell Determinant SOX9 Promotes Lineage Plasticity and Progression in Basal-like Breast Cancer.Cell Rep. 2020 Jun 9;31(10):107742. doi: 10.1016/j.celrep.2020.107742. Cell Rep. 2020. PMID: 32521267 Free PMC article.
-
Evolving insights: how DNA repair pathways impact cancer evolution.Cancer Biol Med. 2020 Nov 15;17(4):805-827. doi: 10.20892/j.issn.2095-3941.2020.0177. Epub 2020 Dec 15. Cancer Biol Med. 2020. PMID: 33299637 Free PMC article. Review.
-
NF-κB role on tumor proliferation, migration, invasion and immune escape.Cancer Gene Ther. 2024 Nov;31(11):1599-1610. doi: 10.1038/s41417-024-00811-6. Epub 2024 Jul 20. Cancer Gene Ther. 2024. PMID: 39033218 Review.
References
-
- Asselin-Labat ML, Sutherland KD, Barker H, Thomas R, Shackleton M, Forrest NC, Hartley L, Robb L, Grosveld FG, van der Wees J, et al. (2007). Gata-3 is an essential regulator of mammary-gland morphogenesis and luminal-cell differentiation. Nat. Cell Biol 9, 201–209. - PubMed
-
- Asselin-Labat ML, Vaillant F, Sheridan JM, Pal B, Wu D, Simpson ER, Yasuda H, Smyth GK, Martin TJ, Lindeman GJ, and Visvader JE (2010). Control of mammary stem cell function by steroid hormone signalling. Nature 465, 798–802. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
