

An evolutionarily conserved mechanism for cAMP elicited axonal regeneration involves direct activation of the dual leucine zipper kinase DLK
- PMID: 27268300
- PMCID: PMC4896747
- DOI: 10.7554/eLife.14048
An evolutionarily conserved mechanism for cAMP elicited axonal regeneration involves direct activation of the dual leucine zipper kinase DLK
Abstract
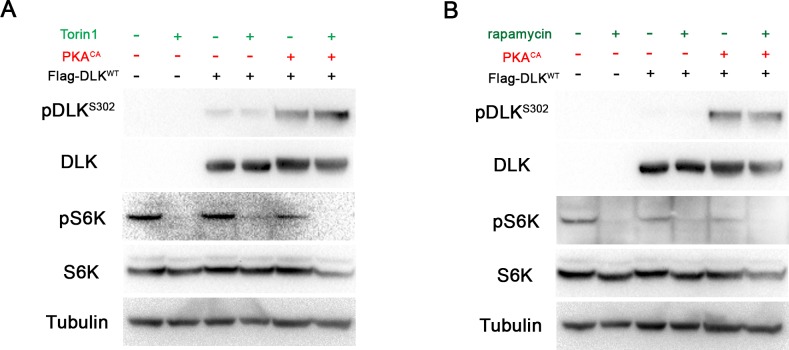
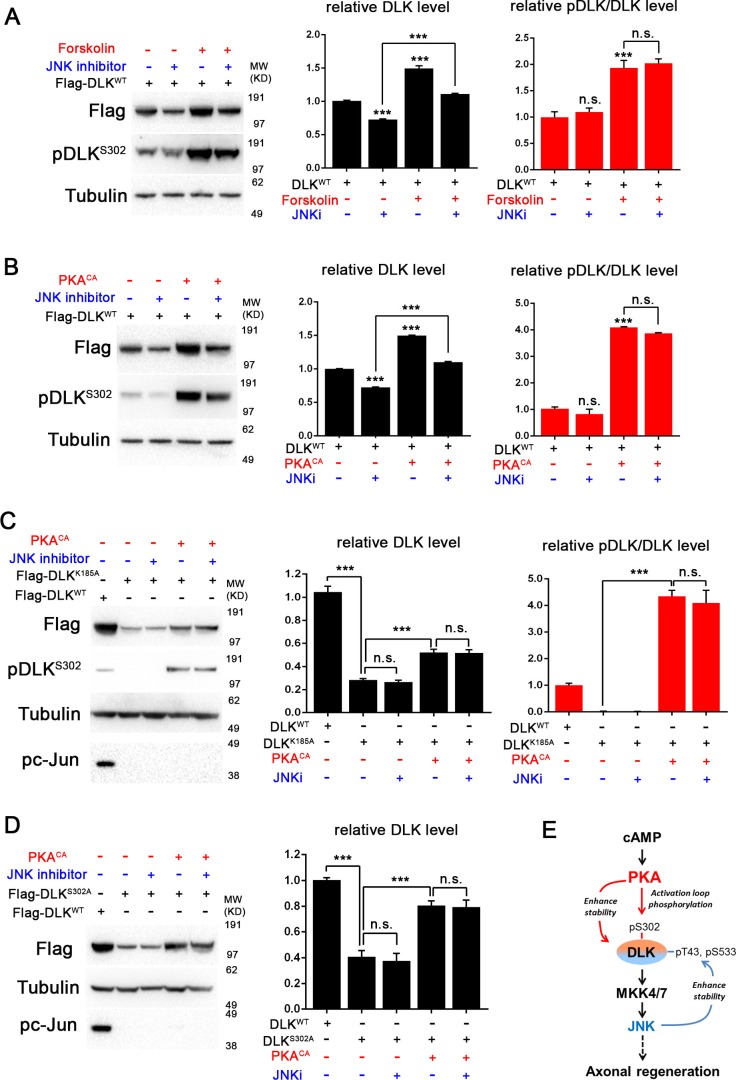
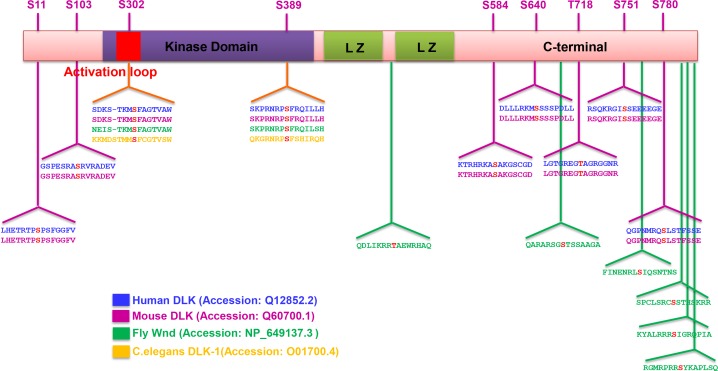
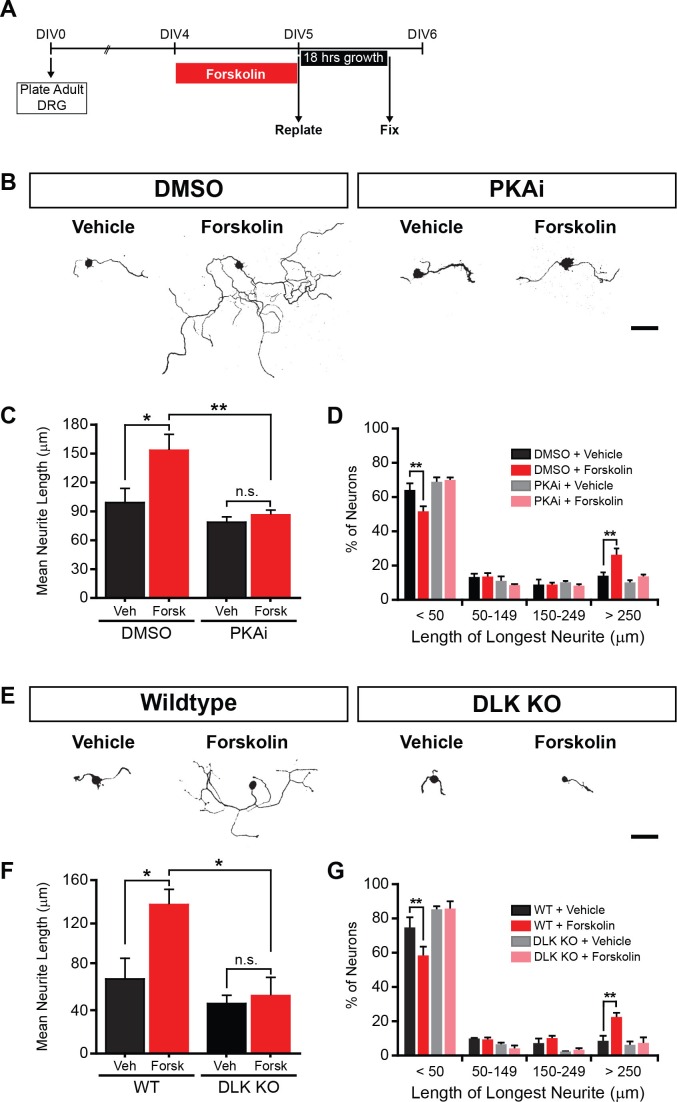
A broadly known method to stimulate the growth potential of axons is to elevate intracellular levels of cAMP, however the cellular pathway(s) that mediate this are not known. Here we identify the Dual Leucine-zipper Kinase (DLK, Wnd in Drosophila) as a critical target and effector of cAMP in injured axons. DLK/Wnd is thought to function as an injury 'sensor', as it becomes activated after axonal damage. Our findings in both Drosophila and mammalian neurons indicate that the cAMP effector kinase PKA is a conserved and direct upstream activator of Wnd/DLK. PKA is required for the induction of Wnd signaling in injured axons, and DLK is essential for the regenerative effects of cAMP in mammalian DRG neurons. These findings link two important mediators of responses to axonal injury, DLK/Wnd and cAMP/PKA, into a unified and evolutionarily conserved molecular pathway for stimulating the regenerative potential of injured axons.
Keywords: D. melanogaster; MAP Kinasae signaling; axonal regeneration; cAMP signaiing; mouse; neuroscience.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Similar articles
-
A Critical Role for DLK and LZK in Axonal Repair in the Mammalian Spinal Cord.J Neurosci. 2022 May 4;42(18):3716-3732. doi: 10.1523/JNEUROSCI.2495-21.2022. Epub 2022 Mar 31. J Neurosci. 2022. PMID: 35361703 Free PMC article.
-
Protein turnover of the Wallenda/DLK kinase regulates a retrograde response to axonal injury.J Cell Biol. 2010 Oct 4;191(1):211-23. doi: 10.1083/jcb.201006039. J Cell Biol. 2010. PMID: 20921142 Free PMC article.
-
Calcium and cyclic AMP promote axonal regeneration in Caenorhabditis elegans and require DLK-1 kinase.J Neurosci. 2010 Mar 3;30(9):3175-83. doi: 10.1523/JNEUROSCI.5464-09.2010. J Neurosci. 2010. PMID: 20203177 Free PMC article.
-
An axonal stress response pathway: degenerative and regenerative signaling by DLK.Curr Opin Neurobiol. 2018 Dec;53:110-119. doi: 10.1016/j.conb.2018.07.002. Epub 2018 Jul 24. Curr Opin Neurobiol. 2018. PMID: 30053694 Free PMC article. Review.
-
The role of cAMP and its downstream targets in neurite growth in the adult nervous system.Neurosci Lett. 2017 Jun 23;652:56-63. doi: 10.1016/j.neulet.2016.12.033. Epub 2016 Dec 15. Neurosci Lett. 2017. PMID: 27989572 Review.
Cited by
-
A cilia-independent function of BBSome mediated by DLK-MAPK signaling in C. elegans photosensation.Dev Cell. 2022 Jun 20;57(12):1545-1557.e4. doi: 10.1016/j.devcel.2022.05.005. Epub 2022 May 31. Dev Cell. 2022. PMID: 35649417 Free PMC article.
-
The Krüppel-Like Factor Gene Target Dusp14 Regulates Axon Growth and Regeneration.Invest Ophthalmol Vis Sci. 2018 Jun 1;59(7):2736-2747. doi: 10.1167/iovs.17-23319. Invest Ophthalmol Vis Sci. 2018. PMID: 29860460 Free PMC article.
-
Role and Interplay of Different Signaling Pathways Involved in Sciatic Nerve Regeneration.J Mol Neurosci. 2024 Nov 12;74(4):108. doi: 10.1007/s12031-024-02286-4. J Mol Neurosci. 2024. PMID: 39531101 Review.
-
Voltage-gated calcium channels act upstream of adenylyl cyclase Ac78C to promote timely initiation of dendrite regeneration.PLoS Genet. 2024 Aug 26;20(8):e1011388. doi: 10.1371/journal.pgen.1011388. eCollection 2024 Aug. PLoS Genet. 2024. PMID: 39186815 Free PMC article.
-
Brief Electrical Stimulation Promotes Recovery after Surgical Repair of Injured Peripheral Nerves.Int J Mol Sci. 2024 Jan 4;25(1):665. doi: 10.3390/ijms25010665. Int J Mol Sci. 2024. PMID: 38203836 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
