

Transcription factor TLX1 controls retinoic acid signaling to ensure spleen development
- PMID: 27214556
- PMCID: PMC4922703
- DOI: 10.1172/JCI82956
Transcription factor TLX1 controls retinoic acid signaling to ensure spleen development
Abstract
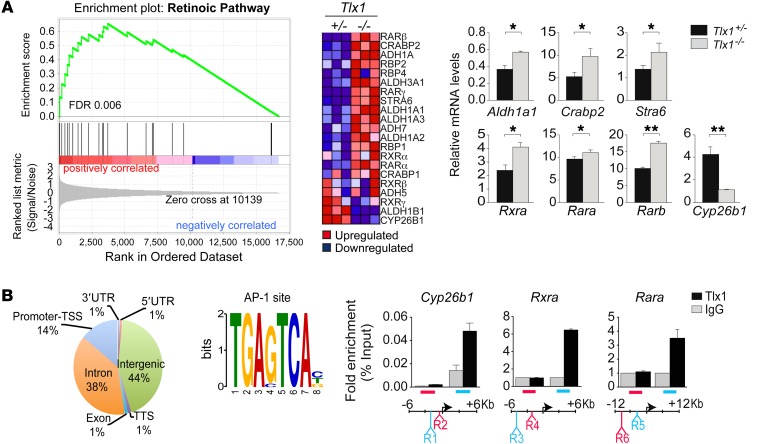
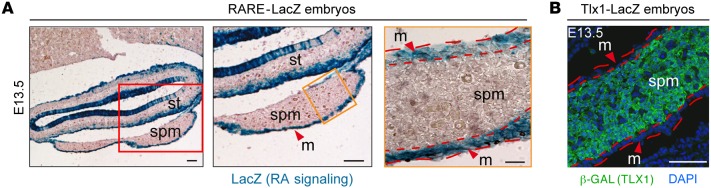
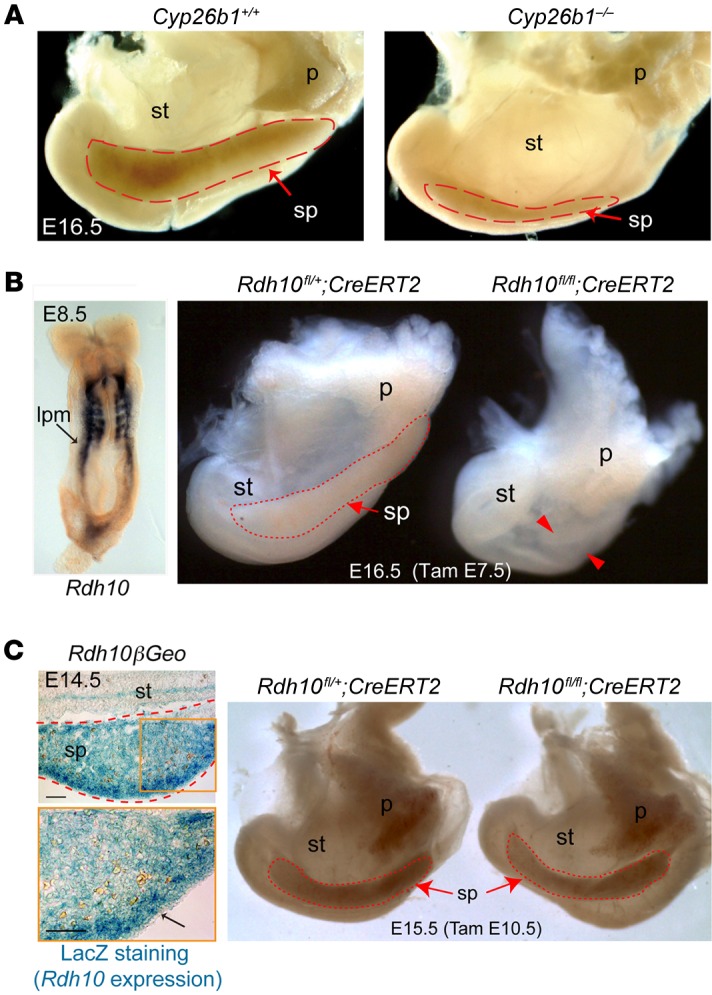
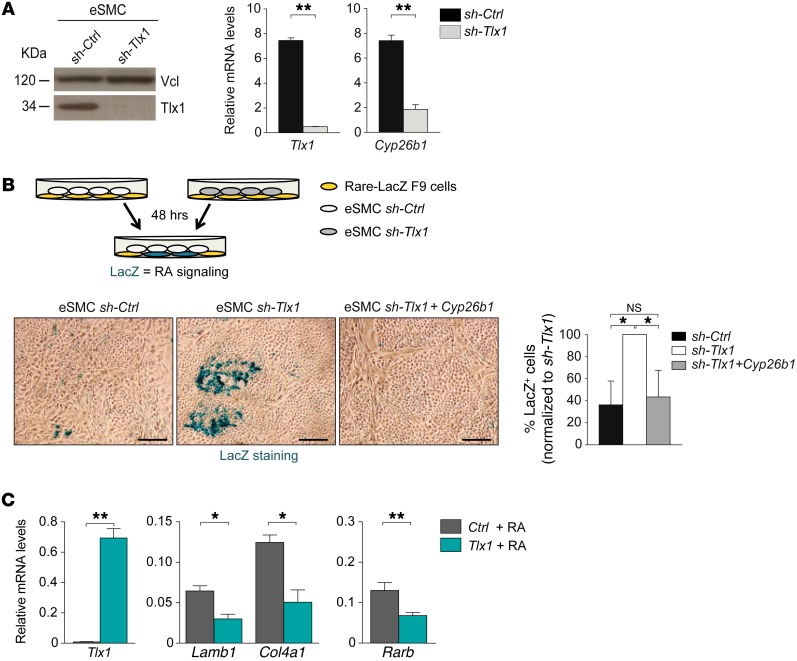
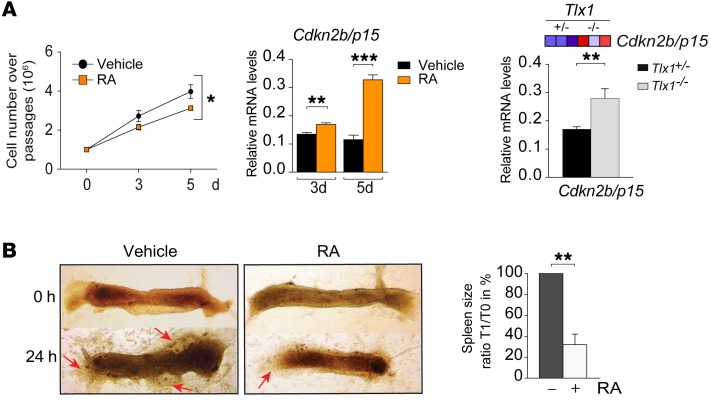
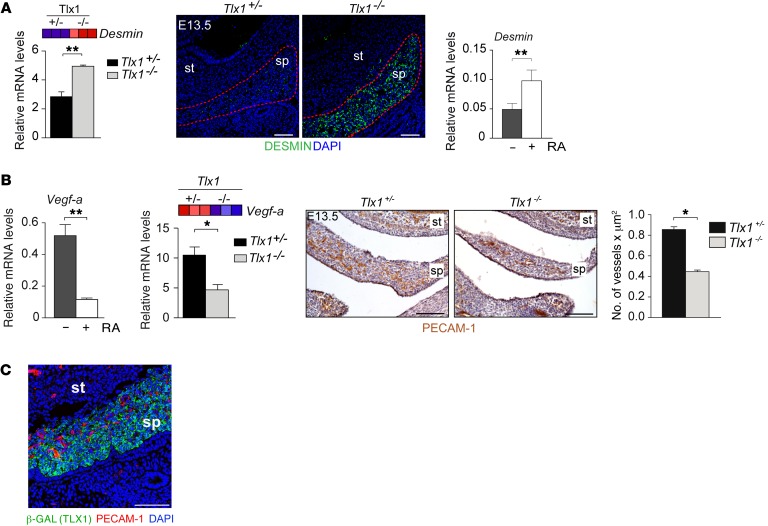
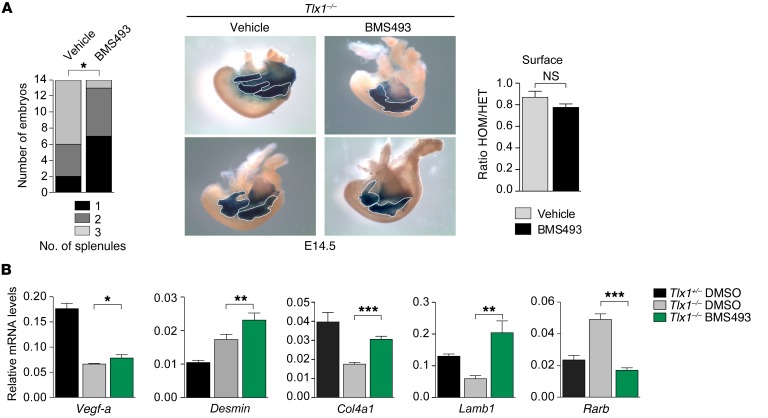
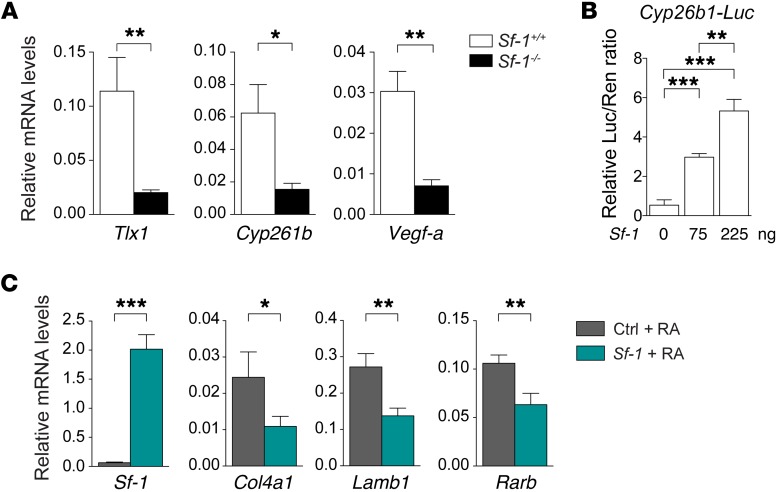
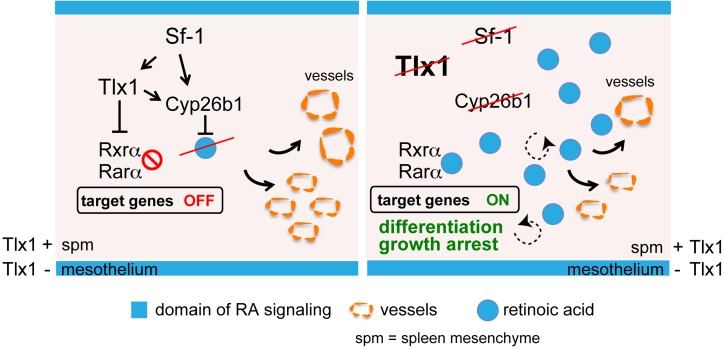
The molecular mechanisms that underlie spleen development and congenital asplenia, a condition linked to increased risk of overwhelming infections, remain largely unknown. The transcription factor TLX1 controls cell fate specification and organ expansion during spleen development, and Tlx1 deletion causes asplenia in mice. Deregulation of TLX1 expression has recently been proposed in the pathogenesis of congenital asplenia in patients carrying mutations of the gene-encoding transcription factor SF-1. Herein, we have shown that TLX1-dependent regulation of retinoic acid (RA) metabolism is critical for spleen organogenesis. In a murine model, loss of Tlx1 during formation of the splenic anlage increased RA signaling by regulating several genes involved in RA metabolism. Uncontrolled RA activity resulted in premature differentiation of mesenchymal cells and reduced vasculogenesis of the splenic primordium. Pharmacological inhibition of RA signaling in Tlx1-deficient animals partially rescued the spleen defect. Finally, spleen growth was impaired in mice lacking either cytochrome P450 26B1 (Cyp26b1), which results in excess RA, or retinol dehydrogenase 10 (Rdh10), which results in RA deficiency. Together, these findings establish TLX1 as a critical regulator of RA metabolism and provide mechanistic insights into the molecular determinants of human congenital asplenia.
Figures
Similar articles
-
Generation of a Tlx1(CreER-Venus) knock-in mouse strain for the study of spleen development.Genesis. 2014 Nov;52(11):916-23. doi: 10.1002/dvg.22829. Epub 2014 Oct 18. Genesis. 2014. PMID: 25283275
-
Congenital asplenia due to a tlx1 mutation reduces resistance to Aeromonas hydrophila infection in zebrafish.Fish Shellfish Immunol. 2019 Dec;95:538-545. doi: 10.1016/j.fsi.2019.10.065. Epub 2019 Oct 31. Fish Shellfish Immunol. 2019. PMID: 31678534
-
Niche-induced extramedullary hematopoiesis in the spleen is regulated by the transcription factor Tlx1.Sci Rep. 2018 May 29;8(1):8308. doi: 10.1038/s41598-018-26693-x. Sci Rep. 2018. PMID: 29844356 Free PMC article.
-
The promise of Hox11+ stem cells of the spleen for treating autoimmune diseases.Horm Metab Res. 2008 Feb;40(2):137-46. doi: 10.1055/s-2007-1022560. Horm Metab Res. 2008. PMID: 18283632 Review.
-
The Cdx1 homeodomain protein: an integrator of posterior signaling in the mouse.Bioessays. 2003 Oct;25(10):971-80. doi: 10.1002/bies.10340. Bioessays. 2003. PMID: 14505364 Review.
Cited by
-
Altered Expression of Candidate Genes in Mayer-Rokitansky-Küster-Hauser Syndrome May Influence Vaginal Keratinocytes Biology: A Focus on Protein Kinase X.Biology (Basel). 2021 May 21;10(6):450. doi: 10.3390/biology10060450. Biology (Basel). 2021. PMID: 34063745 Free PMC article.
-
Expression characteristics and interaction networks of microRNAs in spleen tissues of grass carp (Ctenopharyngodon idella).PLoS One. 2022 Mar 28;17(3):e0266189. doi: 10.1371/journal.pone.0266189. eCollection 2022. PLoS One. 2022. PMID: 35344574 Free PMC article.
-
Retinoid metabolism: new insights.J Mol Endocrinol. 2022 Oct 11;69(4):T37-T49. doi: 10.1530/JME-22-0082. Print 2022 Nov 1. J Mol Endocrinol. 2022. PMID: 35900851 Free PMC article. Review.
-
Splenic stromal niches in homeostasis and immunity.Nat Rev Immunol. 2023 Nov;23(11):705-719. doi: 10.1038/s41577-023-00857-x. Epub 2023 Mar 27. Nat Rev Immunol. 2023. PMID: 36973361 Review.
-
Long Non-Coding RNA BCAR4 Binds to miR-644a and Targets TLX1 to Promote the Progression of Bladder Cancer.Onco Targets Ther. 2020 Mar 24;13:2483-2490. doi: 10.2147/OTT.S232965. eCollection 2020. Onco Targets Ther. 2020. PMID: 32273720 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
