

Medial HOXA genes demarcate haematopoietic stem cell fate during human development
- PMID: 27183470
- PMCID: PMC4981340
- DOI: 10.1038/ncb3354
Medial HOXA genes demarcate haematopoietic stem cell fate during human development
Abstract
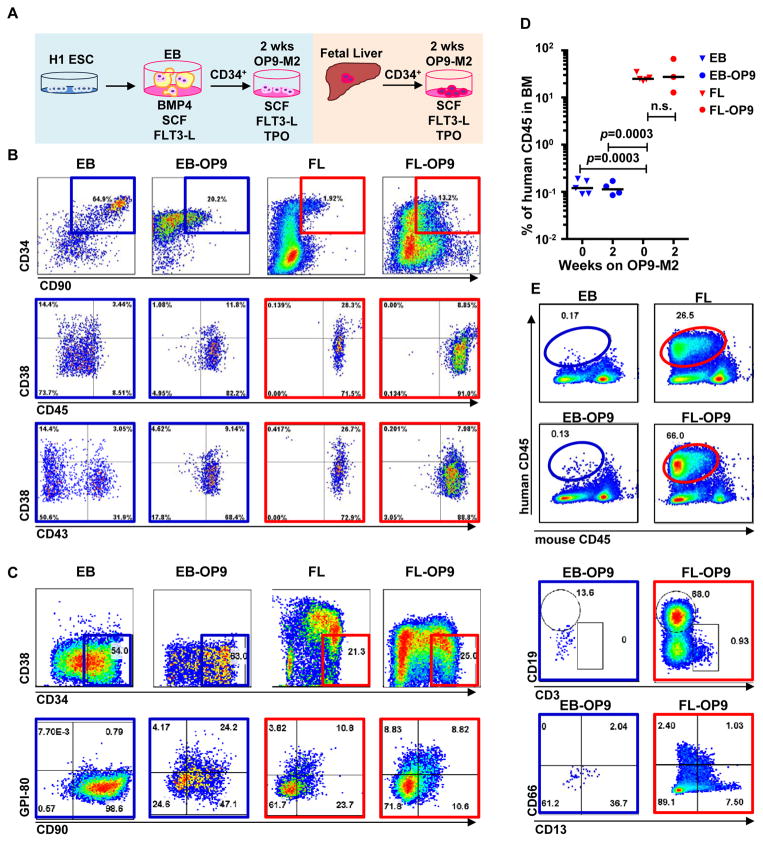
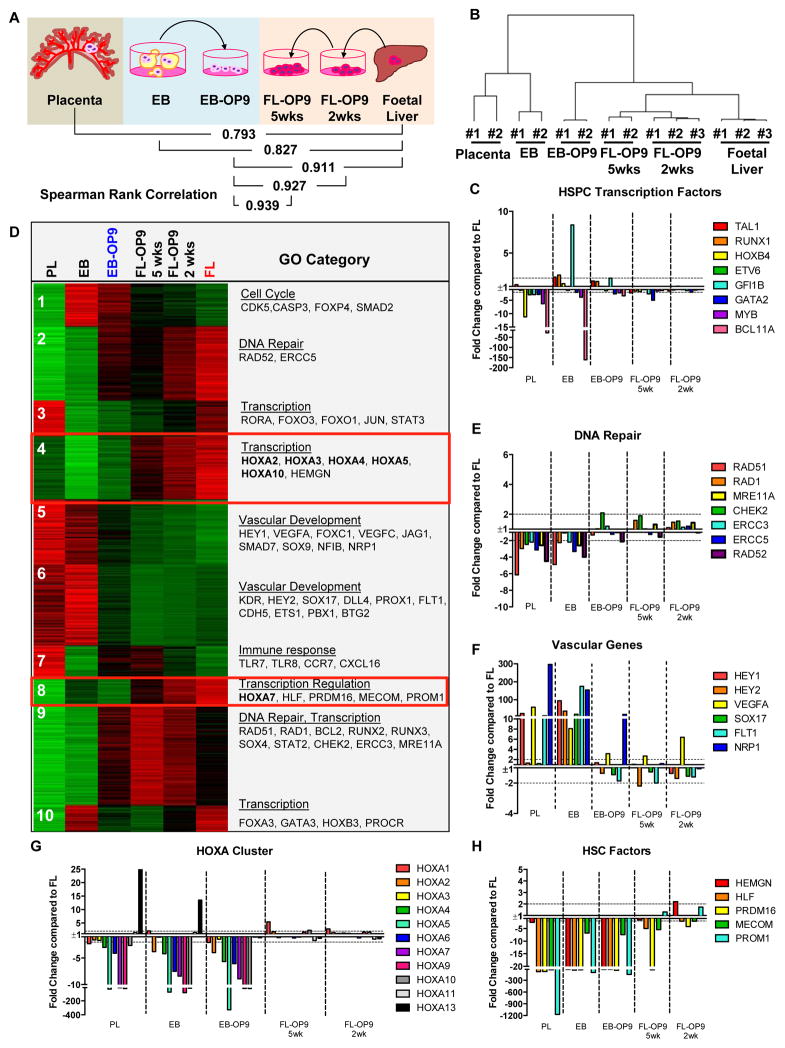
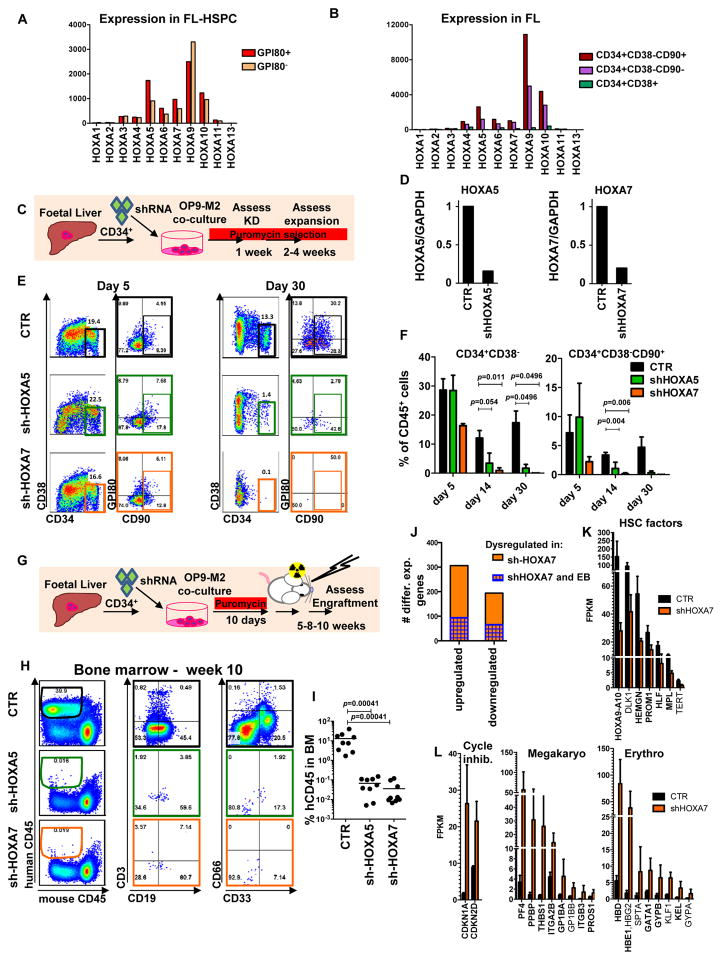
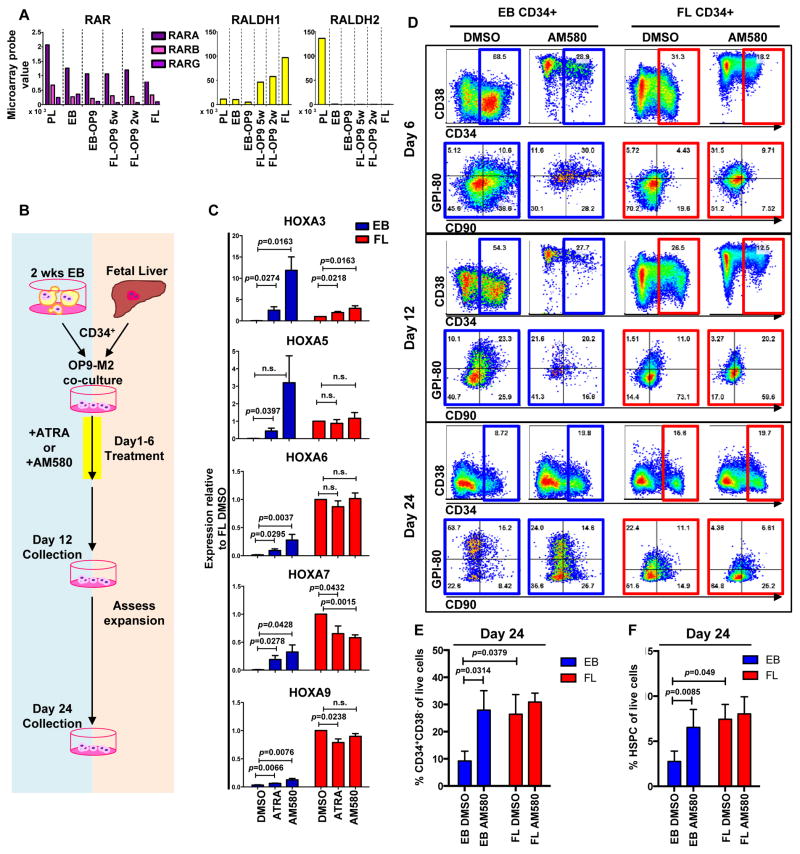
Pluripotent stem cells (PSCs) may provide a potential source of haematopoietic stem/progenitor cells (HSPCs) for transplantation; however, unknown molecular barriers prevent the self-renewal of PSC-HSPCs. Using two-step differentiation, human embryonic stem cells (hESCs) differentiated in vitro into multipotent haematopoietic cells that had the CD34(+)CD38(-/lo)CD90(+)CD45(+)GPI-80(+) fetal liver (FL) HSPC immunophenotype, but exhibited poor expansion potential and engraftment ability. Transcriptome analysis of immunophenotypic hESC-HSPCs revealed that, despite their molecular resemblance to FL-HSPCs, medial HOXA genes remained suppressed. Knockdown of HOXA7 disrupted FL-HSPC function and caused transcriptome dysregulation that resembled hESC-derived progenitors. Overexpression of medial HOXA genes prolonged FL-HSPC maintenance but was insufficient to confer self-renewal to hESC-HSPCs. Stimulation of retinoic acid signalling during endothelial-to-haematopoietic transition induced the HOXA cluster and other HSC/definitive haemogenic endothelium genes, and prolonged HSPC maintenance in culture. Thus, medial HOXA gene expression induced by retinoic acid signalling marks the establishment of the definitive HSPC fate and controls HSPC identity and function.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



Similar articles
-
Extensive immunophenotypic sub-population analysis of StemRegenin1 expanded haematopoietic stem/progenitor cells.Stem Cell Res Ther. 2024 Sep 20;15(1):317. doi: 10.1186/s13287-024-03895-x. Stem Cell Res Ther. 2024. PMID: 39304924 Free PMC article.
-
Endothelial cells provide a niche for placental hematopoietic stem/progenitor cell expansion through broad transcriptomic modification.Stem Cell Res. 2013 Nov;11(3):1074-90. doi: 10.1016/j.scr.2013.07.010. Epub 2013 Aug 9. Stem Cell Res. 2013. PMID: 23978474
-
Three-dimensional co-culture of mesenchymal stromal cells and differentiated osteoblasts on human bio-derived bone scaffolds supports active multi-lineage hematopoiesis in vitro: Functional implication of the biomimetic HSC niche.Int J Mol Med. 2016 Oct;38(4):1141-51. doi: 10.3892/ijmm.2016.2712. Epub 2016 Aug 19. Int J Mol Med. 2016. PMID: 27571775 Free PMC article.
-
Hematopoietic stem cell expansion: challenges and opportunities.Ann N Y Acad Sci. 2012 Aug;1266:138-50. doi: 10.1111/j.1749-6632.2012.06549.x. Ann N Y Acad Sci. 2012. PMID: 22901265 Review.
-
Advances in umbilical cord blood stem cell expansion and clinical translation.Exp Hematol. 2015 Jul;43(7):498-513. doi: 10.1016/j.exphem.2015.04.011. Epub 2015 May 10. Exp Hematol. 2015. PMID: 25970610 Review.
Cited by
-
Inducible Forward Programming of Human Pluripotent Stem Cells to Hemato-endothelial Progenitor Cells with Hematopoietic Progenitor Potential.Stem Cell Reports. 2020 Jan 14;14(1):122-137. doi: 10.1016/j.stemcr.2019.11.005. Epub 2019 Dec 12. Stem Cell Reports. 2020. PMID: 31839543 Free PMC article.
-
Nutrient regulation of development and cell fate decisions.Development. 2023 Oct 15;150(20):dev199961. doi: 10.1242/dev.199961. Epub 2023 Jun 1. Development. 2023. PMID: 37260407 Free PMC article. Review.
-
CTCF boundary remodels chromatin domain and drives aberrant HOX gene transcription in acute myeloid leukemia.Blood. 2018 Aug 23;132(8):837-848. doi: 10.1182/blood-2017-11-814319. Epub 2018 May 14. Blood. 2018. PMID: 29760161 Free PMC article.
-
Activation of the Arterial Program Drives Development of Definitive Hemogenic Endothelium with Lymphoid Potential.Cell Rep. 2018 May 22;23(8):2467-2481. doi: 10.1016/j.celrep.2018.04.092. Cell Rep. 2018. PMID: 29791856 Free PMC article.
-
The hemogenic endothelium: a critical source for the generation of PSC-derived hematopoietic stem and progenitor cells.Cell Mol Life Sci. 2021 May;78(9):4143-4160. doi: 10.1007/s00018-021-03777-y. Epub 2021 Feb 9. Cell Mol Life Sci. 2021. PMID: 33559689 Free PMC article. Review.
References
-
- Bordignon C. Stem-cell therapies for blood diseases. Nature. 2006;441:1100–1102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
