

The Tail Associated Protein of Acinetobacter baumannii Phage ΦAB6 Is the Host Specificity Determinant Possessing Exopolysaccharide Depolymerase Activity
- PMID: 27077375
- PMCID: PMC4831824
- DOI: 10.1371/journal.pone.0153361
The Tail Associated Protein of Acinetobacter baumannii Phage ΦAB6 Is the Host Specificity Determinant Possessing Exopolysaccharide Depolymerase Activity
Abstract
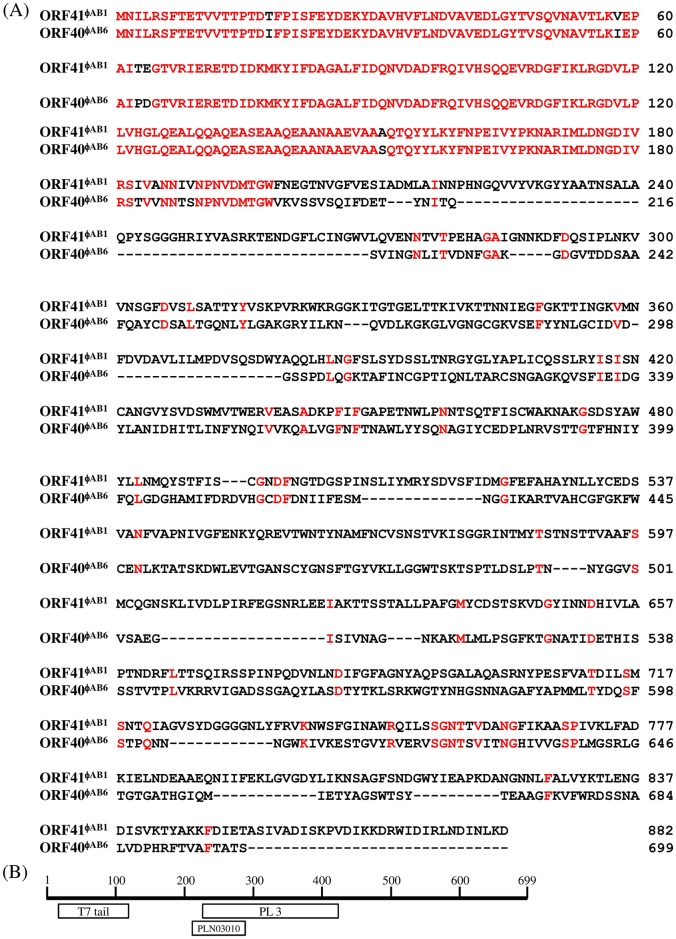
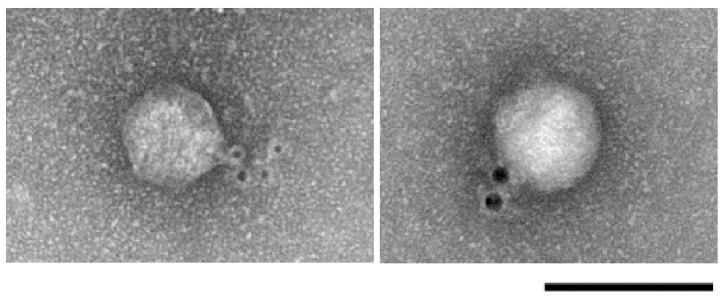
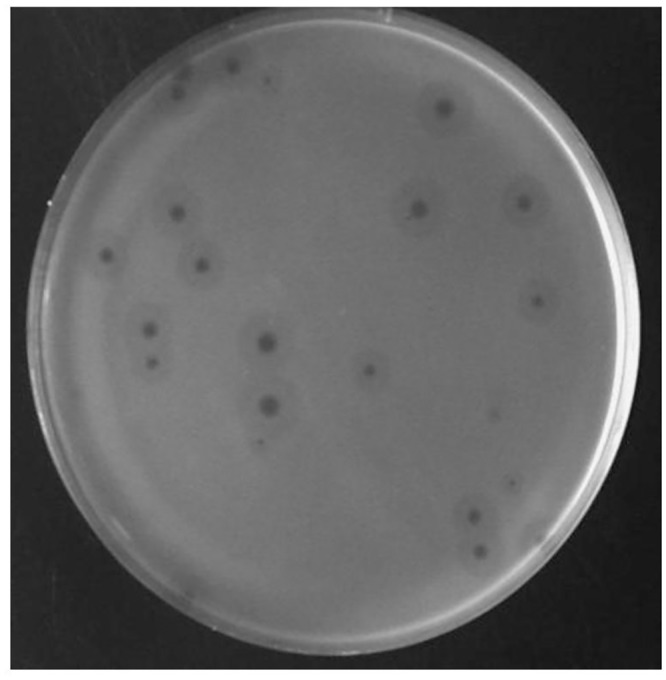
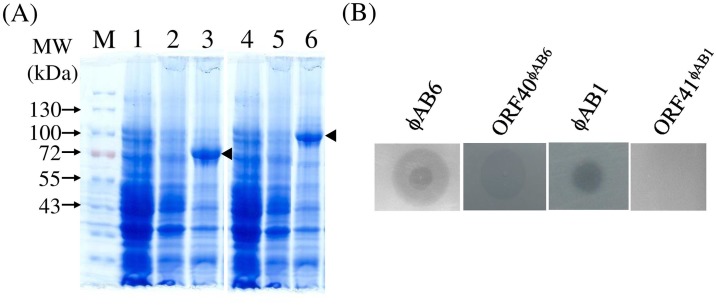
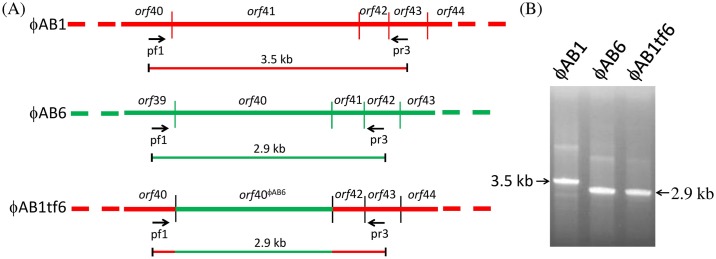
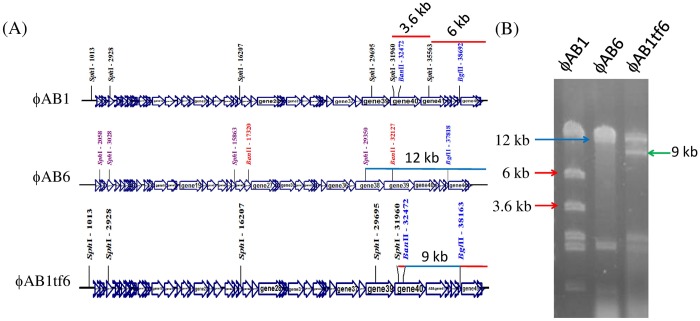
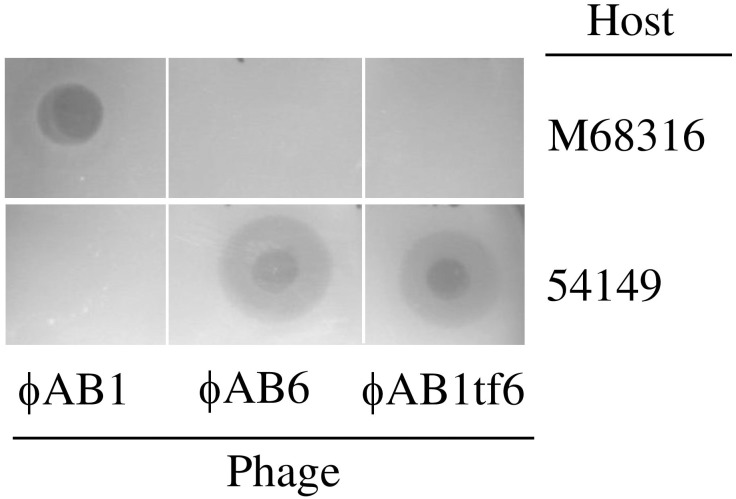
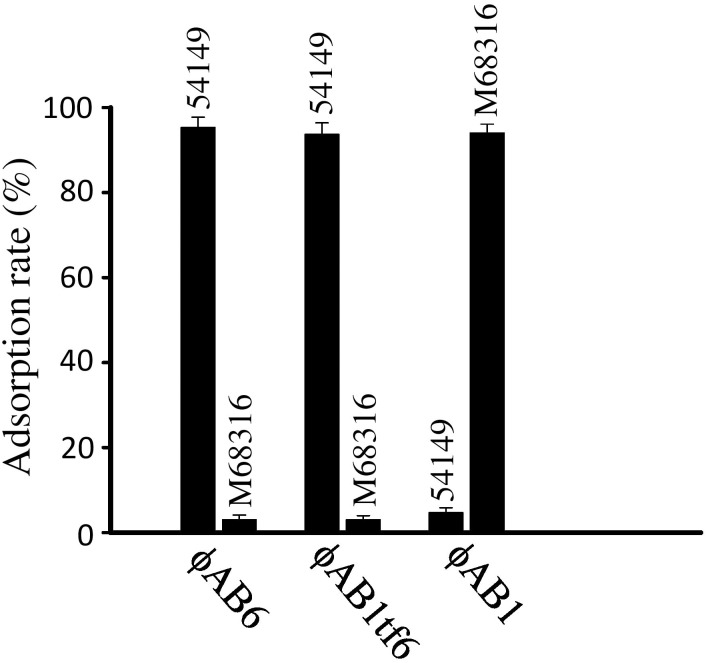
Acinetobacter baumannii is a non-fermenting, gram-negative bacterium. In recent years, the frequency of A. baumannii infections has continued to increase, and multidrug-resistant strains are emerging in hospitalized patients. Therefore, as therapeutic options become limited, the potential of phages as natural antimicrobial agents to control infections is worth reconsidering. In our previous study, we isolated ten virulent double-stranded DNA A. baumannii phages, ϕAB1-9 and ϕAB11, and found that each has a narrow host range. Many reports indicate that receptor-binding protein of phage mediates host recognition; however, understanding of the specific interactions between A. baumannii and phages remains very limited. In this study, host determinants of A. baumannii phages were investigated. Sequence comparison of ϕAB6 and ϕAB1 revealed high degrees of conservation among their genes except the tail fiber protein (ORF41 in ϕAB1 and ORF40 in ϕAB6). Furthermore, we found that ORF40ϕAB6 has polysaccharide depolymerase activity capable of hydrolyzing the A. baumannii exopolysaccharide and is a component of the phage tail apparatus determining host specificity. Thus, the lytic phages and their associated depolymerase not only have potential as alternative therapeutic agents for treating A. baumannii infections but also provide useful and highly specific tools for studying host strain exopolysaccharides and producing glycoconjugate vaccines.
Conflict of interest statement
Figures
Similar articles
-
Functional Analysis and Antivirulence Properties of a New Depolymerase from a Myovirus That Infects Acinetobacter baumannii Capsule K45.J Virol. 2019 Feb 5;93(4):e01163-18. doi: 10.1128/JVI.01163-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30463964 Free PMC article.
-
Novel Acinetobacter baumannii Myovirus TaPaz Encoding Two Tailspike Depolymerases: Characterization and Host-Recognition Strategy.Viruses. 2021 May 25;13(6):978. doi: 10.3390/v13060978. Viruses. 2021. PMID: 34070371 Free PMC article.
-
Genomic and Biochemical Characterization of Acinetobacter Podophage Petty Reveals a Novel Lysis Mechanism and Tail-Associated Depolymerase Activity.J Virol. 2018 Feb 26;92(6):e01064-17. doi: 10.1128/JVI.01064-17. Print 2018 Mar 15. J Virol. 2018. PMID: 29298884 Free PMC article.
-
Specific and Selective Bacteriophages in the Fight against Multidrug-resistant Acinetobacter baumannii.Virol Sin. 2019 Aug;34(4):347-357. doi: 10.1007/s12250-019-00125-0. Epub 2019 May 15. Virol Sin. 2019. PMID: 31093881 Free PMC article. Review.
-
Acinetobacter baumannii Bacteriophage: Progress in Isolation, Genome Sequencing, Preclinical Research, and Clinical Application.Curr Microbiol. 2023 Apr 30;80(6):199. doi: 10.1007/s00284-023-03295-z. Curr Microbiol. 2023. PMID: 37120784 Free PMC article. Review.
Cited by
-
Phage Digestion of a Bacterial Capsule Imparts Resistance to Two Antibiotic Agents.Microorganisms. 2021 Apr 10;9(4):794. doi: 10.3390/microorganisms9040794. Microorganisms. 2021. PMID: 33920206 Free PMC article.
-
Isolation and Characterization of a Phapecoctavirus Infecting Multidrug-Resistant Acinetobacter baumannii in A549 Alveolar Epithelial Cells.Viruses. 2022 Nov 19;14(11):2561. doi: 10.3390/v14112561. Viruses. 2022. PMID: 36423170 Free PMC article.
-
Expansion of the Plaquing Host Range and Improvement of the Absorption Rate of a T5-like Salmonella Phage by Altering the Long Tail Fibers.Appl Environ Microbiol. 2022 Sep 13;88(17):e0089522. doi: 10.1128/aem.00895-22. Epub 2022 Aug 15. Appl Environ Microbiol. 2022. PMID: 35969059 Free PMC article.
-
Comparative Analysis of 37 Acinetobacter Bacteriophages.Viruses. 2017 Dec 24;10(1):5. doi: 10.3390/v10010005. Viruses. 2017. PMID: 29295549 Free PMC article.
-
Therapeutic Application of Bacteriophage PHB02 and Its Putative Depolymerase Against Pasteurella multocida Capsular Type A in Mice.Front Microbiol. 2018 Aug 7;9:1678. doi: 10.3389/fmicb.2018.01678. eCollection 2018. Front Microbiol. 2018. PMID: 30131774 Free PMC article.
References
-
- Duplessis M, Moineau S. Identification of a genetic determinant responsible for host specificity in Streptococcus thermophilus bacteriophages. Mol Microbiol. 2001; 41: 325–336. - PubMed
-
- Rakhuba DV, Kolomiets EI, Dey ES, Novik GI. Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol J Microbiol. 2010; 59: 145–155. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
