

CCR5 interaction with HIV-1 Env contributes to Env-induced depletion of CD4 T cells in vitro and in vivo
- PMID: 27026376
- PMCID: PMC4812640
- DOI: 10.1186/s12977-016-0255-z
CCR5 interaction with HIV-1 Env contributes to Env-induced depletion of CD4 T cells in vitro and in vivo
Abstract
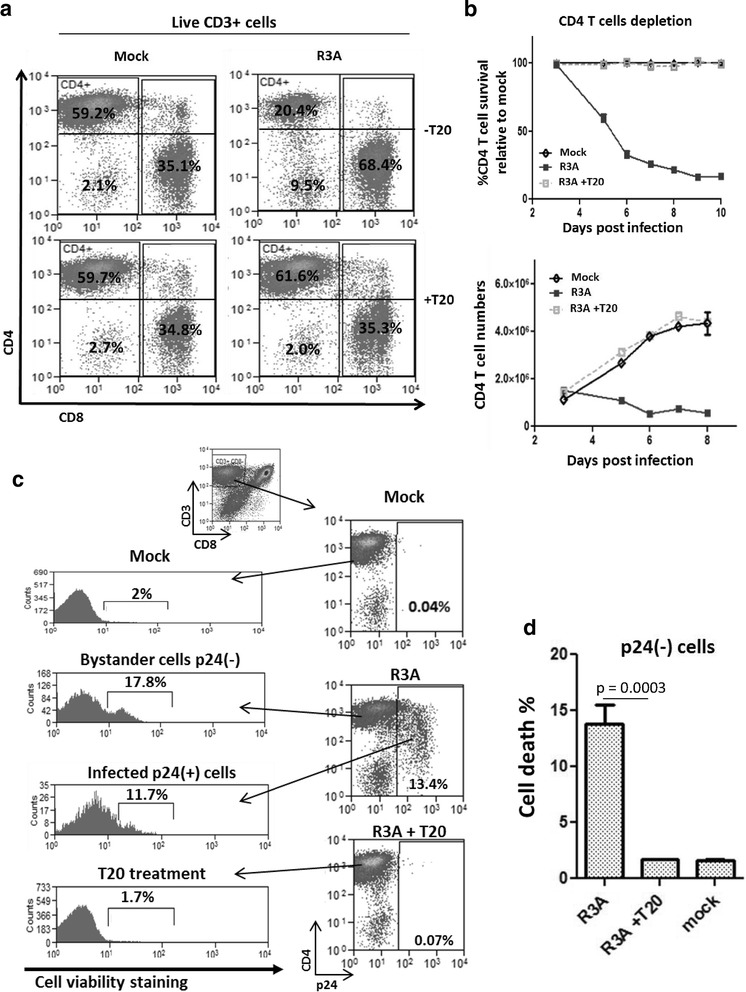
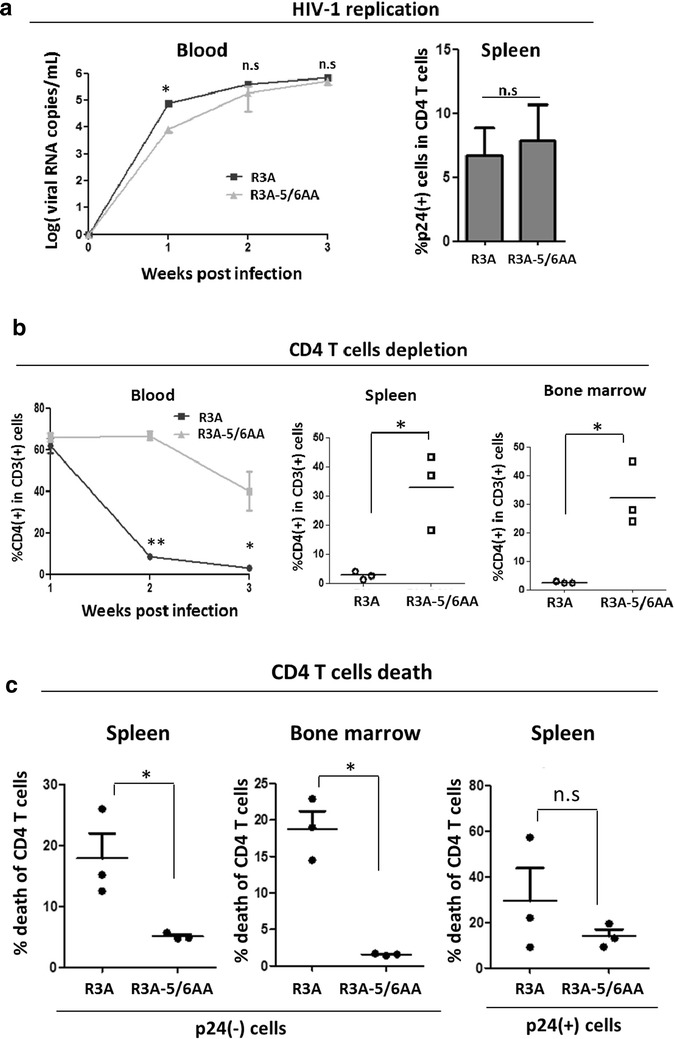
Background: CD4 T cell depletion during HIV-1 infection is associated with AIDS disease progression, and the HIV-1 Env protein plays an important role in the process. Together with CXCR4, CCR5 is one of the two co-receptors that interact with Env during virus entry, but the role of CCR5 in Env-induced pathogenesis is not clearly defined. We have investigated CD4 T cell depletion mechanisms caused by the Env of a highly pathogenic CXCR4/CCR5 dual-tropic HIV-1 isolate R3A.
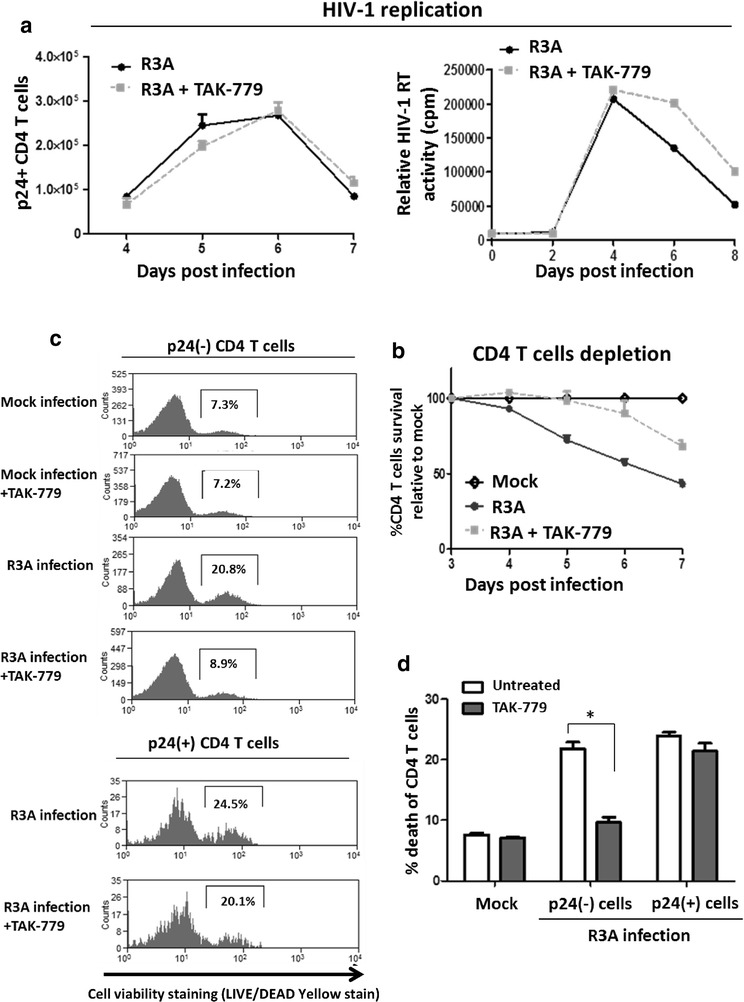
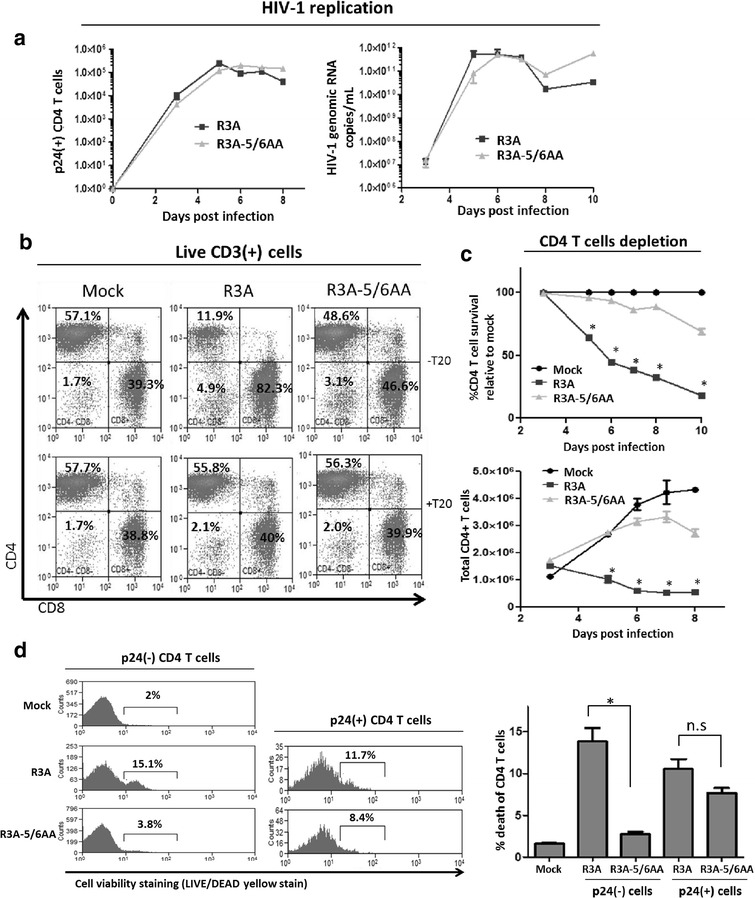
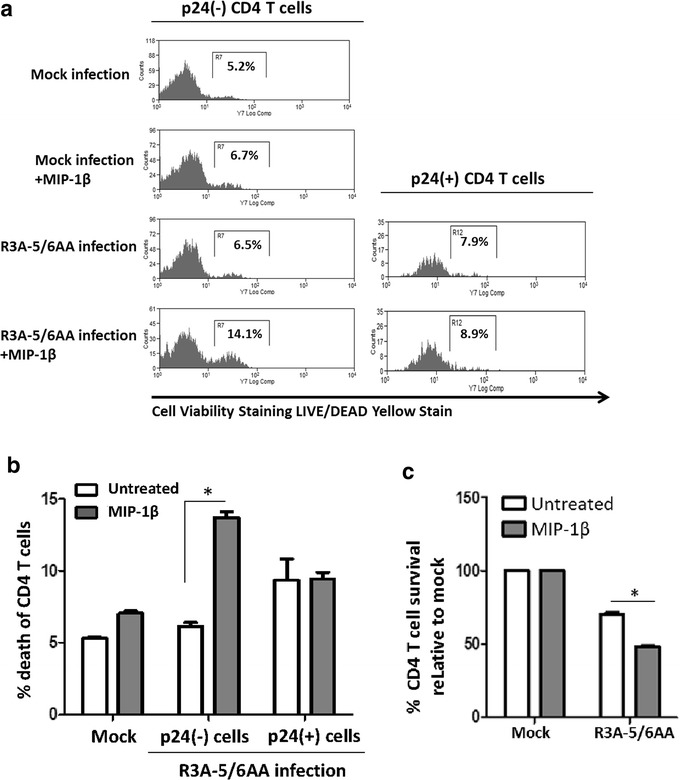
Results: We report here that R3A infection induced depletion of both infected and uninfected "bystander" CD4 T cells, and treatment with CCR5 antagonist TAK-779 inhibited R3A-induced bystander CD4 T cell depletion without affecting virus replication. To further define the role of Env-CCR5 interaction, we utilized an Env-mutant of R3A, termed R3A-5/6AA, which has lost CCR5 binding capability. Importantly, R3A-5/6AA replicated to the same level as wild type R3A by using CXCR4 for viral infection. We found the loss of CCR5 interaction resulted in a significant reduction of bystander CD4 T cells death during R3A-5/6AA infection, whereas stimulation of CCR5 with MIP1-β increased bystander pathogenesis induced by R3A-5/6AA. We confirmed our findings using a humanized mouse model, where we observed similarly reduced pathogenicity of the mutant R3A-5/6AA in various lymphoid organs in vivo.
Conclusion: We provide the first evidence that shows CCR5 interaction with a dual-tropic HIV-1 Env played a significant role in Env-induced depletion of CD4 T cells.
Keywords: Bystander CD4 T cells; CCR5; HIV-1 Env; HIV-1 pathogenesis; Humanized mice.
Figures
Similar articles
-
Apoptosis of bystander T cells induced by human immunodeficiency virus type 1 with increased envelope/receptor affinity and coreceptor binding site exposure.J Virol. 2004 May;78(9):4541-51. doi: 10.1128/jvi.78.9.4541-4551.2004. J Virol. 2004. PMID: 15078935 Free PMC article.
-
Critical roles for Akt kinase in controlling HIV envelope-mediated depletion of CD4 T cells.Retrovirology. 2013 Jun 6;10:60. doi: 10.1186/1742-4690-10-60. Retrovirology. 2013. PMID: 23742646 Free PMC article.
-
Host and Viral Factors in HIV-Mediated Bystander Apoptosis.Viruses. 2017 Aug 22;9(8):237. doi: 10.3390/v9080237. Viruses. 2017. PMID: 28829402 Free PMC article. Review.
-
HIV-1 Env and Nef Cooperatively Contribute to Plasmacytoid Dendritic Cell Activation via CD4-Dependent Mechanisms.J Virol. 2015 Aug;89(15):7604-11. doi: 10.1128/JVI.00695-15. Epub 2015 May 13. J Virol. 2015. PMID: 25972534 Free PMC article.
-
The SIV Envelope Glycoprotein, Viral Tropism, and Pathogenesis: Novel Insights from Nonhuman Primate Models of AIDS.Curr HIV Res. 2018;16(1):29-40. doi: 10.2174/1570162X15666171124123116. Curr HIV Res. 2018. PMID: 29173176 Review.
Cited by
-
Multiplexed shRNA-miRs as a candidate for anti HIV-1 therapy: strategies, challenges, and future potential.J Genet Eng Biotechnol. 2022 Dec 28;20(1):172. doi: 10.1186/s43141-022-00451-z. J Genet Eng Biotechnol. 2022. PMID: 36576612 Free PMC article. Review.
-
Lymphopenia Caused by Virus Infections and the Mechanisms Beyond.Viruses. 2021 Sep 20;13(9):1876. doi: 10.3390/v13091876. Viruses. 2021. PMID: 34578457 Free PMC article. Review.
-
The Interplay of HIV and Autophagy in Early Infection.Front Microbiol. 2021 Apr 28;12:661446. doi: 10.3389/fmicb.2021.661446. eCollection 2021. Front Microbiol. 2021. PMID: 33995324 Free PMC article. Review.
-
Differential Pathogenicity of SHIV KB9 and 89.6 Env Correlates with Bystander Apoptosis Induction in CD4+ T cells.Viruses. 2019 Oct 1;11(10):911. doi: 10.3390/v11100911. Viruses. 2019. PMID: 31581579 Free PMC article.
-
Sexual Dimorphism in Innate Immunity.Clin Rev Allergy Immunol. 2019 Jun;56(3):308-321. doi: 10.1007/s12016-017-8648-x. Clin Rev Allergy Immunol. 2019. PMID: 28963611 Review.
References
-
- Hazenberg MD, Hamann D, Schuitemaker H, Miedema F. T cell depletion in HIV-1 infection : how CD4+ T cells go out of stock. Cell Death Dis. 2000;1:285–9. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
