

Citrullination-acetylation interplay guides E2F-1 activity during the inflammatory response
- PMID: 26989780
- PMCID: PMC4788482
- DOI: 10.1126/sciadv.1501257
Citrullination-acetylation interplay guides E2F-1 activity during the inflammatory response
Abstract
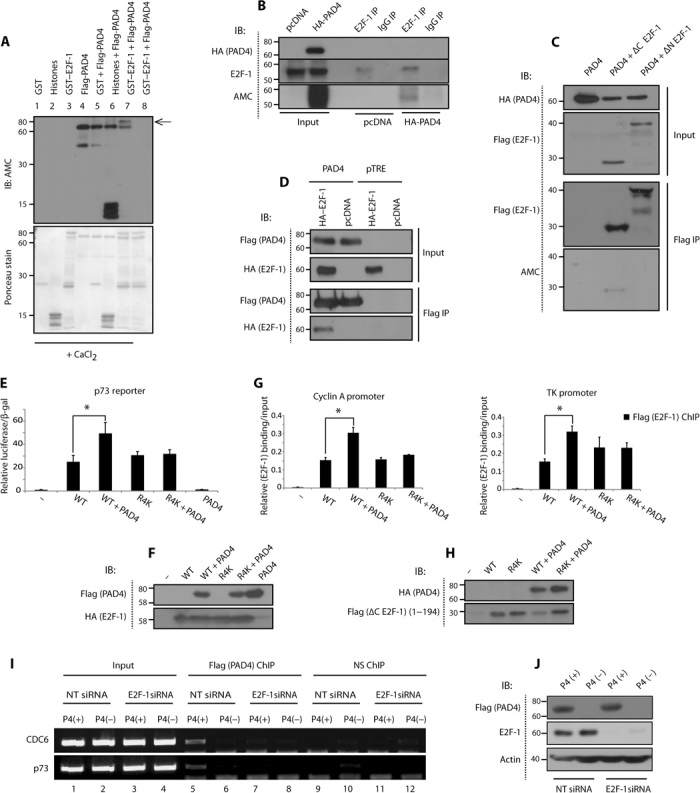
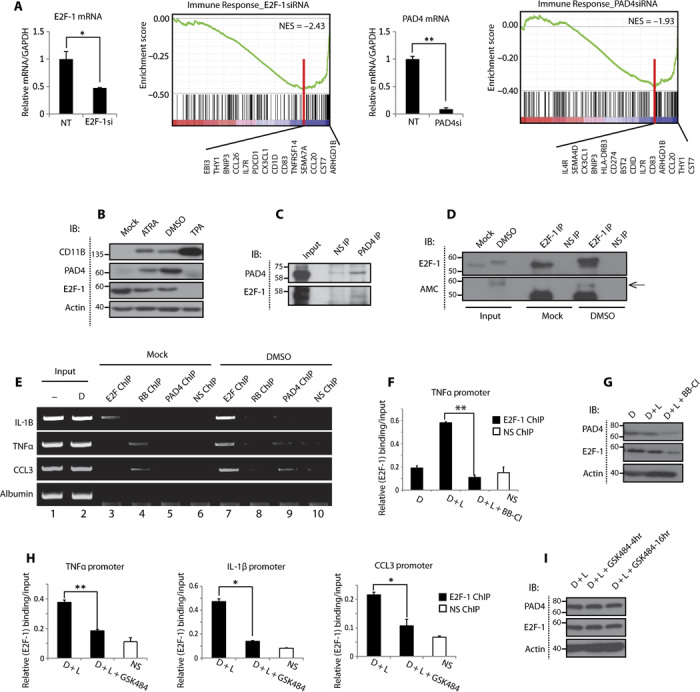
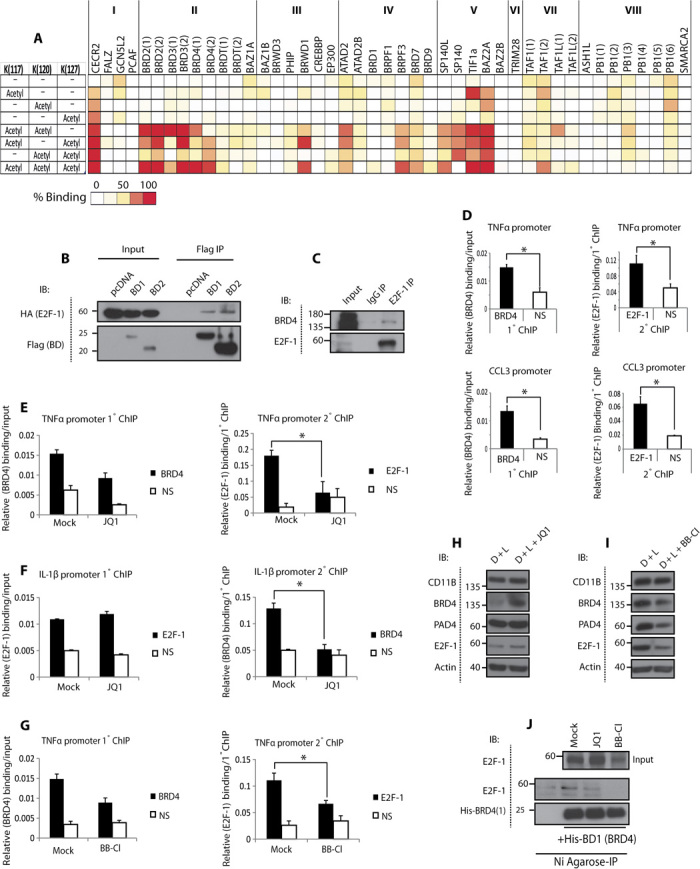
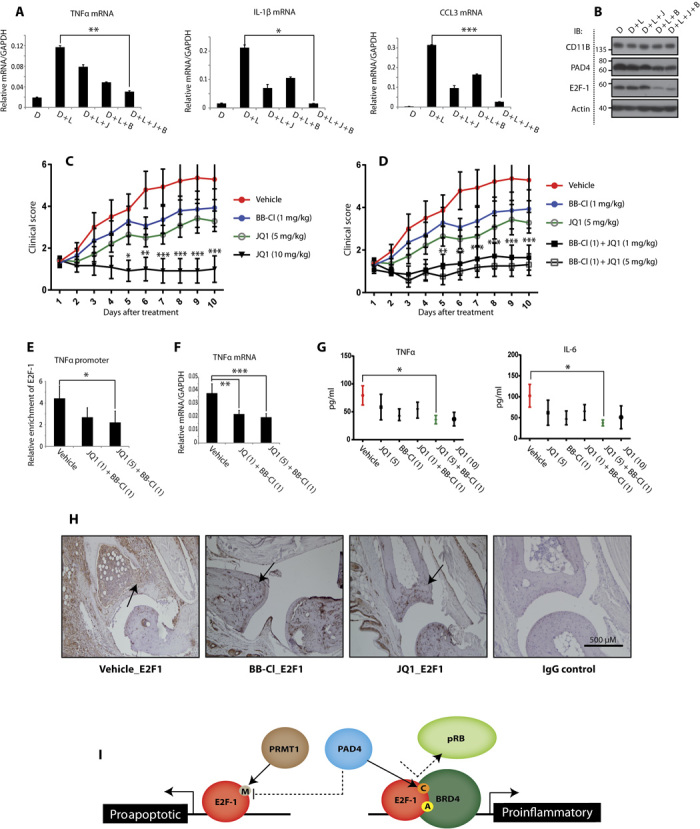
Peptidyl arginine deiminase 4 (PAD4) is a nuclear enzyme that converts arginine residues to citrulline. Although increasingly implicated in inflammatory disease and cancer, the mechanism of action of PAD4 and its functionally relevant pathways remains unclear. E2F transcription factors are a family of master regulators that coordinate gene expression during cellular proliferation and diverse cell fates. We show that E2F-1 is citrullinated by PAD4 in inflammatory cells. Citrullination of E2F-1 assists its chromatin association, specifically to cytokine genes in granulocyte cells. Mechanistically, citrullination augments binding of the BET (bromodomain and extra-terminal domain) family bromodomain reader BRD4 (bromodomain-containing protein 4) to an acetylated domain in E2F-1, and PAD4 and BRD4 coexist with E2F-1 on cytokine gene promoters. Accordingly, the combined inhibition of PAD4 and BRD4 disrupts the chromatin-bound complex and suppresses cytokine gene expression. In the murine collagen-induced arthritis model, chromatin-bound E2F-1 in inflammatory cells and consequent cytokine expression are diminished upon small-molecule inhibition of PAD4 and BRD4, and the combined treatment is clinically efficacious in preventing disease progression. Our results shed light on a new transcription-based mechanism that mediates the inflammatory effect of PAD4 and establish the interplay between citrullination and acetylation in the control of E2F-1 as a regulatory interface for driving inflammatory gene expression.
Keywords: BRD4; E2F-1; PAD4; cancer; citrullination; immune response; inflammation.
Figures
Similar articles
-
PAD4-dependent citrullination of nuclear translocation of GSK3β promotes colorectal cancer progression via the degradation of nuclear CDKN1A.Neoplasia. 2022 Nov;33:100835. doi: 10.1016/j.neo.2022.100835. Epub 2022 Sep 13. Neoplasia. 2022. PMID: 36113195 Free PMC article.
-
Citrullination of inhibitor of growth 4 (ING4) by peptidylarginine deminase 4 (PAD4) disrupts the interaction between ING4 and p53.J Biol Chem. 2011 May 13;286(19):17069-78. doi: 10.1074/jbc.M111.230961. Epub 2011 Mar 22. J Biol Chem. 2011. PMID: 21454715 Free PMC article.
-
Peptidylarginine deiminase 2 is required for tumor necrosis factor alpha-induced citrullination and arthritis, but not neutrophil extracellular trap formation.J Autoimmun. 2017 Jun;80:39-47. doi: 10.1016/j.jaut.2017.01.006. Epub 2017 Feb 7. J Autoimmun. 2017. PMID: 28188029 Free PMC article.
-
Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis.Biochim Biophys Acta. 2013 Oct;1829(10):1126-35. doi: 10.1016/j.bbagrm.2013.07.003. Epub 2013 Jul 13. Biochim Biophys Acta. 2013. PMID: 23860259 Free PMC article. Review.
-
PAD4: pathophysiology, current therapeutics and future perspective in rheumatoid arthritis.Expert Opin Ther Targets. 2017 Apr;21(4):433-447. doi: 10.1080/14728222.2017.1294160. Epub 2017 Feb 22. Expert Opin Ther Targets. 2017. PMID: 28281906 Review.
Cited by
-
Candida albicans-Induced NETosis Is Independent of Peptidylarginine Deiminase 4.Front Immunol. 2018 Jul 9;9:1573. doi: 10.3389/fimmu.2018.01573. eCollection 2018. Front Immunol. 2018. PMID: 30038623 Free PMC article.
-
Reciprocal regulation of Th2 and Th17 cells by PAD2-mediated citrullination.JCI Insight. 2019 Nov 14;4(22):e129687. doi: 10.1172/jci.insight.129687. JCI Insight. 2019. PMID: 31723060 Free PMC article.
-
Development of Activity-Based Proteomic Probes for Protein Citrullination.Curr Top Microbiol Immunol. 2019;420:233-251. doi: 10.1007/82_2018_132. Curr Top Microbiol Immunol. 2019. PMID: 30203394 Free PMC article. Review.
-
Cutting Edge: Protein Arginine Deiminase 2 and 4 Regulate NLRP3 Inflammasome-Dependent IL-1β Maturation and ASC Speck Formation in Macrophages.J Immunol. 2019 Aug 15;203(4):795-800. doi: 10.4049/jimmunol.1800720. Epub 2019 Jul 10. J Immunol. 2019. PMID: 31292215 Free PMC article.
-
BRD4 regulates cellular senescence in gastric cancer cells via E2F/miR-106b/p21 axis.Cell Death Dis. 2018 Feb 12;9(2):203. doi: 10.1038/s41419-017-0181-6. Cell Death Dis. 2018. PMID: 29434197 Free PMC article.
References
-
- Christophorou M. A., Castelo-Branco G., Halley-Stott R. P., Oliveira C. S., Loos R., Radzisheuskaya A., Mowen K. A., Bertone P., Silva J. C. R., Zernicka-Goetz M., Nielsen M. L., Gurdon J. B., Kouzarides T., Citrullination regulates pluripotency and histone H1 binding to chromatin. Nature 507, 104–108 (2014). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
