

Dynamics of epigenetic regulation at the single-cell level
- PMID: 26912859
- PMCID: PMC5108652
- DOI: 10.1126/science.aab2956
Dynamics of epigenetic regulation at the single-cell level
Abstract
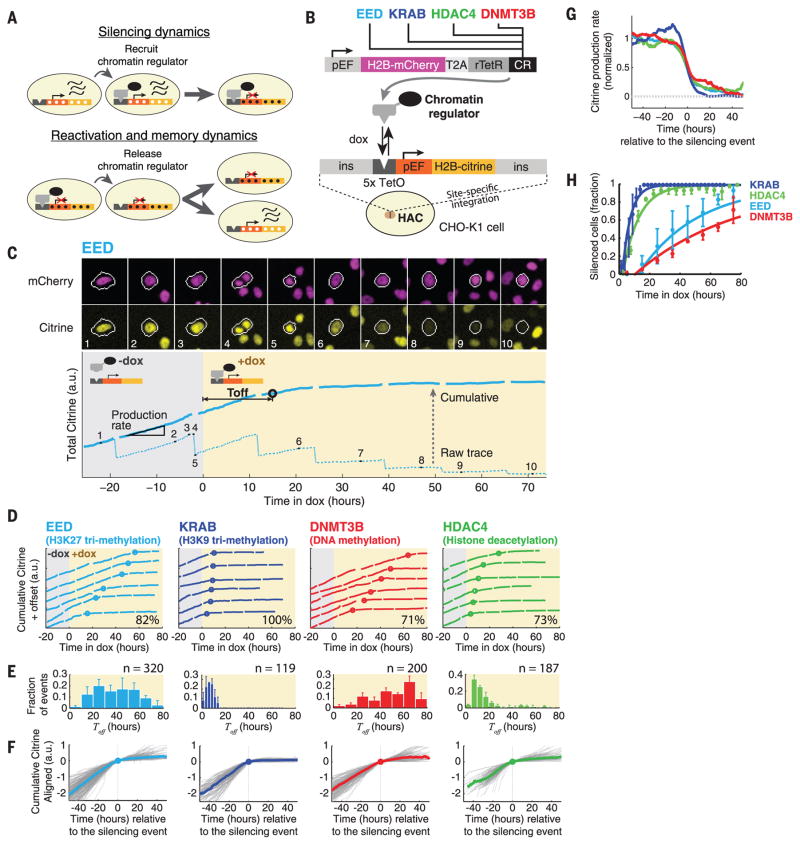
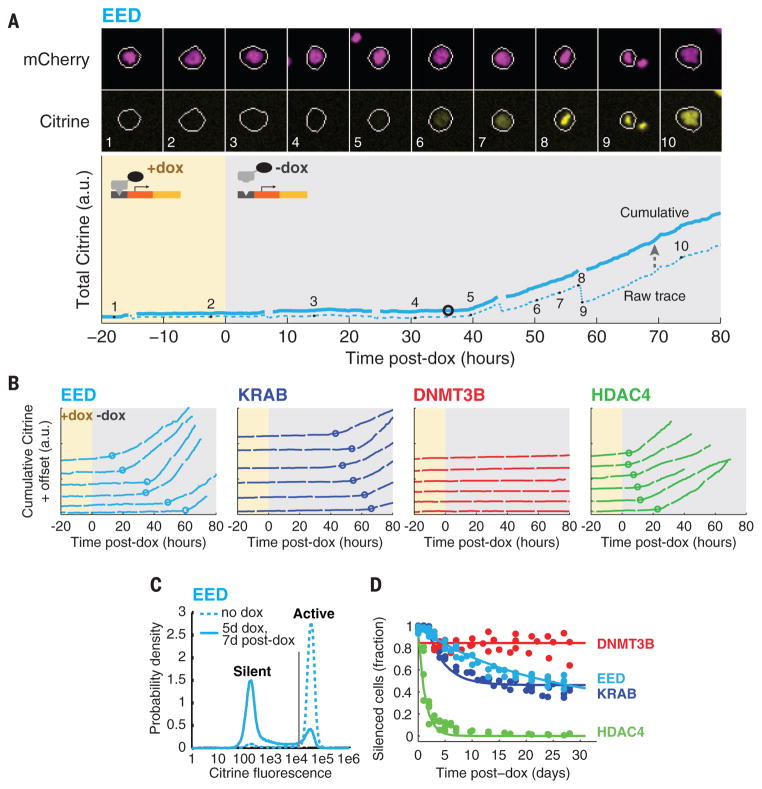
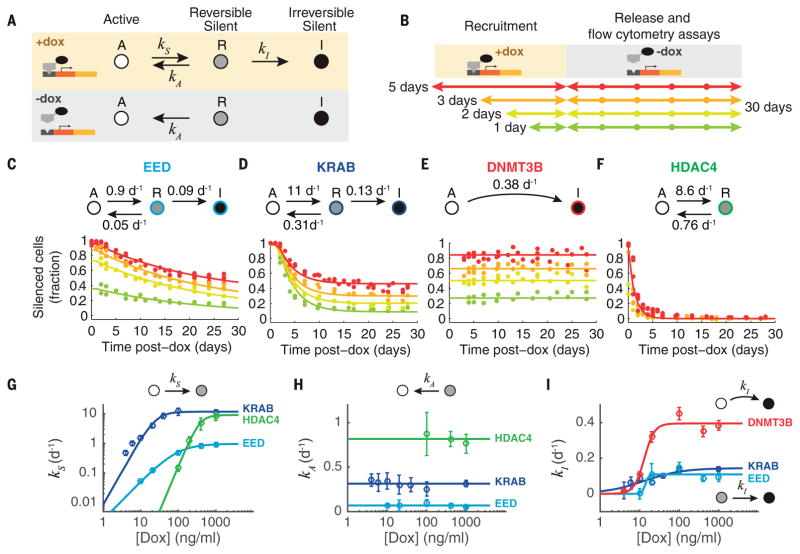
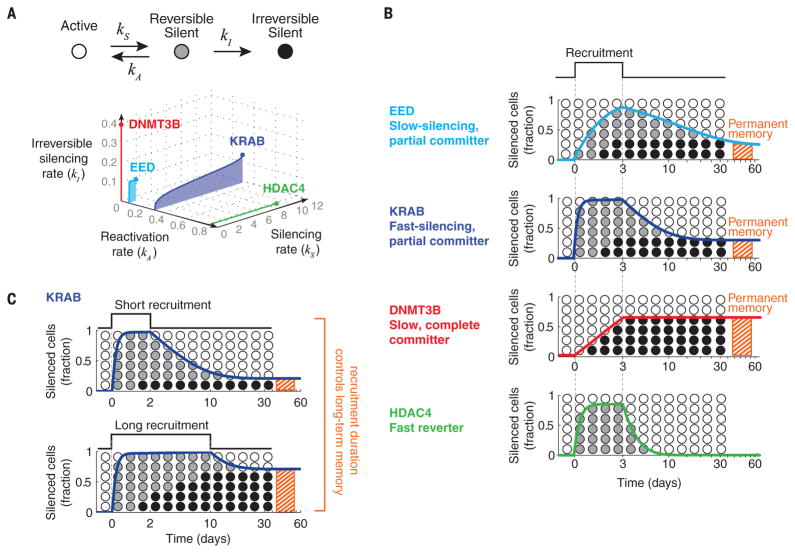
Chromatin regulators play a major role in establishing and maintaining gene expression states. Yet how they control gene expression in single cells, quantitatively and over time, remains unclear. We used time-lapse microscopy to analyze the dynamic effects of four silencers associated with diverse modifications: DNA methylation, histone deacetylation, and histone methylation. For all regulators, silencing and reactivation occurred in all-or-none events, enabling the regulators to modulate the fraction of cells silenced rather than the amount of gene expression. These dynamics could be described by a three-state model involving stochastic transitions between active, reversibly silent, and irreversibly silent states. Through their individual transition rates, these regulators operate over different time scales and generate distinct types of epigenetic memory. Our results provide a framework for understanding and engineering mammalian chromatin regulation and epigenetic memory.
Copyright © 2016, American Association for the Advancement of Science.
Figures
Comment in
-
Epigenetics: A three-state model for epigenetic silencing.Nat Rev Genet. 2016 Apr;17(4):192-3. doi: 10.1038/nrg.2016.19. Epub 2016 Feb 22. Nat Rev Genet. 2016. PMID: 26900023 No abstract available.
-
MOLECULAR BIOLOGY. A unifying model of epigenetic regulation.Science. 2016 Feb 12;351(6274):661-2. doi: 10.1126/science.aaf1647. Science. 2016. PMID: 26912843 Free PMC article. No abstract available.
Similar articles
-
Reactivation of silenced genes and transcriptional therapy.Cytogenet Genome Res. 2003;100(1-4):56-64. doi: 10.1159/000072838. Cytogenet Genome Res. 2003. PMID: 14526164 Review.
-
Methylation-mediated silencing of TMS1/ASC is accompanied by histone hypoacetylation and CpG island-localized changes in chromatin architecture.J Biol Chem. 2002 Feb 15;277(7):4951-8. doi: 10.1074/jbc.M109809200. Epub 2001 Dec 3. J Biol Chem. 2002. PMID: 11733524
-
DNA methyltransferase 1 knock down induces gene expression by a mechanism independent of DNA methylation and histone deacetylation.J Biol Chem. 2004 Jul 2;279(27):27915-27. doi: 10.1074/jbc.M312823200. Epub 2004 Apr 15. J Biol Chem. 2004. PMID: 15087453
-
DNA methylation and histone deacetylation in the control of gene expression: basic biochemistry to human development and disease.Gene Expr. 2000;9(1-2):63-75. doi: 10.3727/000000001783992731. Gene Expr. 2000. PMID: 11097425 Free PMC article. Review.
-
Effect of dynamic DNA methylation and histone acetylation on cPouV expression in differentiation of chick embryonic germ cells.Stem Cells Dev. 2013 Oct 15;22(20):2725-35. doi: 10.1089/scd.2013.0046. Epub 2013 Jul 17. Stem Cells Dev. 2013. PMID: 23750509 Free PMC article.
Cited by
-
DNA hypomethylation promotes the expression of CASPASE-4 which exacerbates inflammation and amyloid-β deposition in Alzheimer's disease.Alzheimers Res Ther. 2024 Feb 8;16(1):29. doi: 10.1186/s13195-024-01390-2. Alzheimers Res Ther. 2024. PMID: 38326859 Free PMC article.
-
Tunable, division-independent control of gene activation timing by a polycomb switch.Cell Rep. 2021 Mar 23;34(12):108888. doi: 10.1016/j.celrep.2021.108888. Cell Rep. 2021. PMID: 33761349 Free PMC article.
-
Single-cell image analysis to explore cell-to-cell heterogeneity in isogenic populations.Cell Syst. 2021 Jun 16;12(6):608-621. doi: 10.1016/j.cels.2021.05.010. Cell Syst. 2021. PMID: 34139168 Free PMC article. Review.
-
Transcription factors perform a 2-step search of the nucleus.Genetics. 2022 Sep 30;222(2):iyac111. doi: 10.1093/genetics/iyac111. Genetics. 2022. PMID: 35939561 Free PMC article.
-
The Promises and Challenges of Toxico-Epigenomics: Environmental Chemicals and Their Impacts on the Epigenome.Environ Health Perspect. 2020 Jan;128(1):15001. doi: 10.1289/EHP6104. Epub 2020 Jan 17. Environ Health Perspect. 2020. PMID: 31950866 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
