

APOBEC-induced mutations in human cancers are strongly enriched on the lagging DNA strand during replication
- PMID: 26755635
- PMCID: PMC4728370
- DOI: 10.1101/gr.197046.115
APOBEC-induced mutations in human cancers are strongly enriched on the lagging DNA strand during replication
Abstract
APOBEC3A and APOBEC3B, cytidine deaminases of the APOBEC family, are among the main factors causing mutations in human cancers. APOBEC deaminates cytosines in single-stranded DNA (ssDNA). A fraction of the APOBEC-induced mutations occur as clusters ("kataegis") in single-stranded DNA produced during repair of double-stranded breaks (DSBs). However, the properties of the remaining 87% of nonclustered APOBEC-induced mutations, the source and the genomic distribution of the ssDNA where they occur, are largely unknown. By analyzing genomic and exomic cancer databases, we show that >33% of dispersed APOBEC-induced mutations occur on the lagging strand during DNA replication, thus unraveling the major source of ssDNA targeted by APOBEC in cancer. Although methylated cytosine is generally more mutation-prone than nonmethylated cytosine, we report that methylation reduces the rate of APOBEC-induced mutations by a factor of roughly two. Finally, we show that in cancers with extensive APOBEC-induced mutagenesis, there is almost no increase in mutation rates in late replicating regions (contrary to other cancers). Because late-replicating regions are depleted in exons, this results in a 1.3-fold higher fraction of mutations residing within exons in such cancers. This study provides novel insight into the APOBEC-induced mutagenesis and describes the peculiarity of the mutational processes in cancers with the signature of APOBEC-induced mutations.
© 2016 Seplyarskiy et al.; Published by Cold Spring Harbor Laboratory Press.
Figures

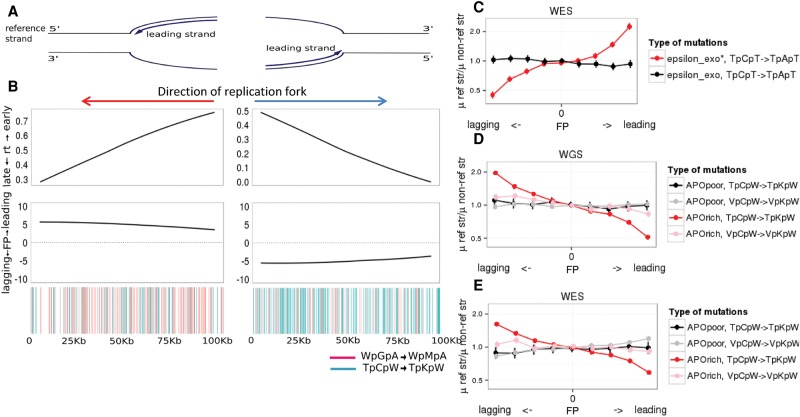

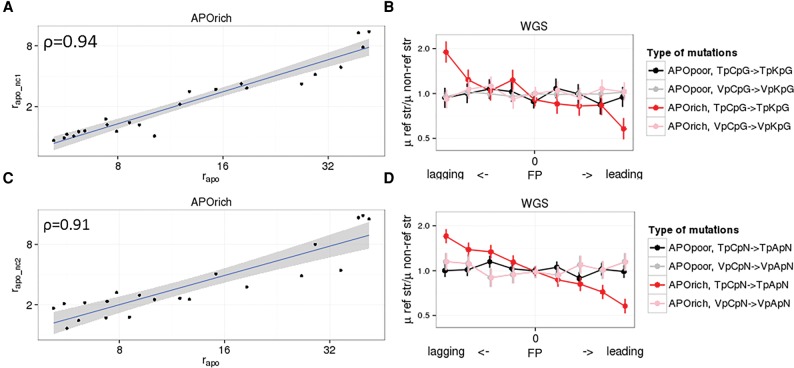
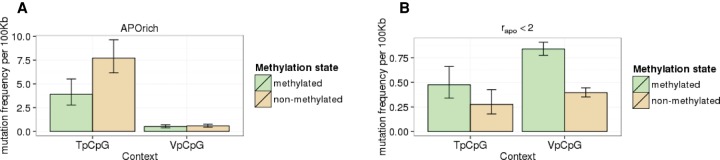
Similar articles
-
An APOBEC3A hypermutation signature is distinguishable from the signature of background mutagenesis by APOBEC3B in human cancers.Nat Genet. 2015 Sep;47(9):1067-72. doi: 10.1038/ng.3378. Epub 2015 Aug 10. Nat Genet. 2015. PMID: 26258849 Free PMC article.
-
APOBEC3A and APOBEC3B Preferentially Deaminate the Lagging Strand Template during DNA Replication.Cell Rep. 2016 Feb 16;14(6):1273-1282. doi: 10.1016/j.celrep.2016.01.021. Epub 2016 Jan 28. Cell Rep. 2016. PMID: 26832400 Free PMC article.
-
Strand-biased cytosine deamination at the replication fork causes cytosine to thymine mutations in Escherichia coli.Proc Natl Acad Sci U S A. 2016 Feb 23;113(8):2176-81. doi: 10.1073/pnas.1522325113. Epub 2016 Feb 2. Proc Natl Acad Sci U S A. 2016. PMID: 26839411 Free PMC article.
-
Error-free versus mutagenic processing of genomic uracil--relevance to cancer.DNA Repair (Amst). 2014 Jul;19:38-47. doi: 10.1016/j.dnarep.2014.03.028. Epub 2014 Apr 18. DNA Repair (Amst). 2014. PMID: 24746924 Review.
-
Breaking bad: The mutagenic effect of DNA repair.DNA Repair (Amst). 2015 Aug;32:43-51. doi: 10.1016/j.dnarep.2015.04.012. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 26073774 Free PMC article. Review.
Cited by
-
HPV16 and HPV18 Genome Structure, Expression, and Post-Transcriptional Regulation.Int J Mol Sci. 2022 Apr 29;23(9):4943. doi: 10.3390/ijms23094943. Int J Mol Sci. 2022. PMID: 35563334 Free PMC article. Review.
-
RNA Editors, Cofactors, and mRNA Targets: An Overview of the C-to-U RNA Editing Machinery and Its Implication in Human Disease.Genes (Basel). 2018 Dec 27;10(1):13. doi: 10.3390/genes10010013. Genes (Basel). 2018. PMID: 30591678 Free PMC article. Review.
-
Analysis of gene expression and mutation data points on contribution of transcription to the mutagenesis by APOBEC enzymes.NAR Cancer. 2021 Jul 2;3(3):zcab025. doi: 10.1093/narcan/zcab025. eCollection 2021 Sep. NAR Cancer. 2021. PMID: 34316712 Free PMC article.
-
Differentiation signals induce APOBEC3A expression via GRHL3 in squamous epithelia and squamous cell carcinoma.EMBO J. 2025 Jan;44(1):1-29. doi: 10.1038/s44318-024-00298-9. Epub 2024 Nov 15. EMBO J. 2025. PMID: 39548236 Free PMC article.
-
Regulation, functional impact, and therapeutic targeting of APOBEC3A in cancer.DNA Repair (Amst). 2024 Sep;141:103734. doi: 10.1016/j.dnarep.2024.103734. Epub 2024 Jul 20. DNA Repair (Amst). 2024. PMID: 39047499 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources