

Myeloid differentiation architecture of leukocyte transcriptome dynamics in perceived social isolation
- PMID: 26598672
- PMCID: PMC4679065
- DOI: 10.1073/pnas.1514249112
Myeloid differentiation architecture of leukocyte transcriptome dynamics in perceived social isolation
Abstract
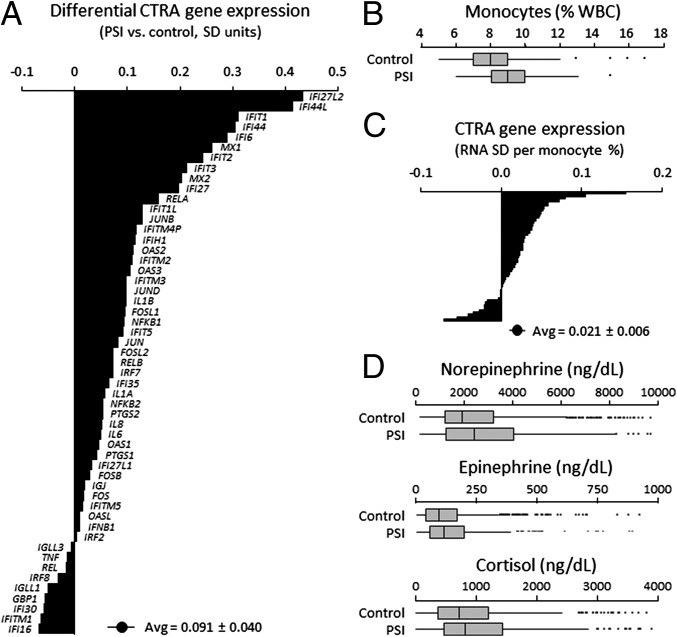
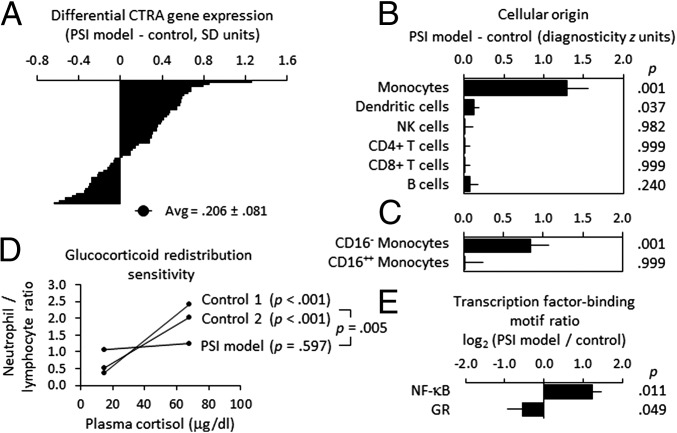
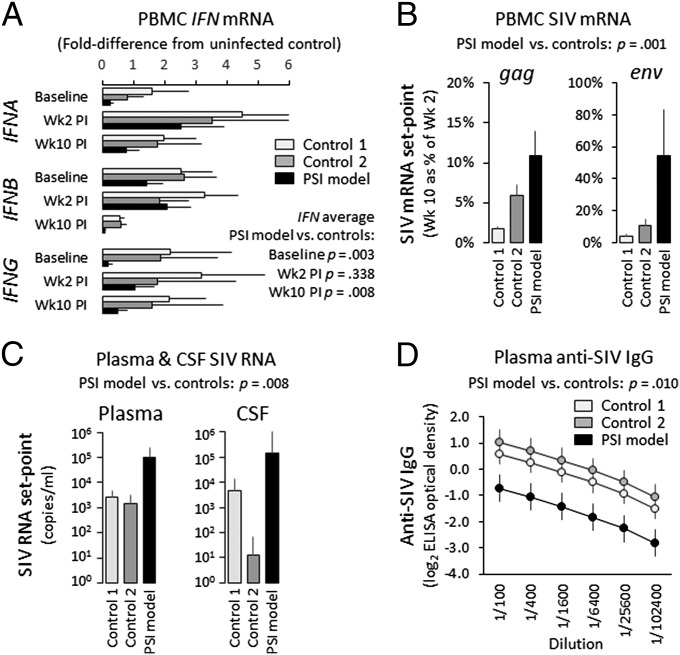
To define the cellular mechanisms of up-regulated inflammatory gene expression and down-regulated antiviral response in people experiencing perceived social isolation (loneliness), we conducted integrative analyses of leukocyte gene regulation in humans and rhesus macaques. Five longitudinal leukocyte transcriptome surveys in 141 older adults showed up-regulation of the sympathetic nervous system (SNS), monocyte population expansion, and up-regulation of the leukocyte conserved transcriptional response to adversity (CTRA). Mechanistic analyses in a macaque model of perceived social isolation confirmed CTRA activation and identified selective up-regulation of the CD14(++)/CD16(-) classical monocyte transcriptome, functional glucocorticoid desensitization, down-regulation of Type I and II interferons, and impaired response to infection by simian immunodeficiency virus (SIV). These analyses identify neuroendocrine-related alterations in myeloid cell population dynamics as a key mediator of CTRA transcriptome skewing, which may both propagate perceived social isolation and contribute to its associated health risks.
Keywords: health; inflammation; loneliness; social genomics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Loneliness, eudaimonia, and the human conserved transcriptional response to adversity.Psychoneuroendocrinology. 2015 Dec;62:11-7. doi: 10.1016/j.psyneuen.2015.07.001. Epub 2015 Jul 8. Psychoneuroendocrinology. 2015. PMID: 26246388 Free PMC article.
-
Social stress up-regulates inflammatory gene expression in the leukocyte transcriptome via β-adrenergic induction of myelopoiesis.Proc Natl Acad Sci U S A. 2013 Oct 8;110(41):16574-9. doi: 10.1073/pnas.1310655110. Epub 2013 Sep 23. Proc Natl Acad Sci U S A. 2013. PMID: 24062448 Free PMC article.
-
Social regulation of the lymph node transcriptome in rhesus macaques (Macaca mulatta).Psychoneuroendocrinology. 2017 Feb;76:107-113. doi: 10.1016/j.psyneuen.2016.10.029. Epub 2016 Nov 14. Psychoneuroendocrinology. 2017. PMID: 27902946 Free PMC article.
-
Implication of Spiritual Network Support System in Epigenomic Modulation and Health Trajectory.Int J Environ Res Public Health. 2019 Oct 25;16(21):4123. doi: 10.3390/ijerph16214123. Int J Environ Res Public Health. 2019. PMID: 31717711 Free PMC article. Review.
-
Human social genomics.PLoS Genet. 2014 Aug 28;10(8):e1004601. doi: 10.1371/journal.pgen.1004601. eCollection 2014 Aug. PLoS Genet. 2014. PMID: 25166010 Free PMC article. Review.
Cited by
-
The impact of social relationships on the risk of stroke and post-stroke mortality: a systematic review and meta-analysis.BMC Public Health. 2024 Sep 4;24(1):2403. doi: 10.1186/s12889-024-19835-6. BMC Public Health. 2024. PMID: 39232685 Free PMC article.
-
Loneliness and the persistence of fear: Perceived social isolation reduces evaluative fear extinction.PLoS One. 2024 Aug 19;19(8):e0303895. doi: 10.1371/journal.pone.0303895. eCollection 2024. PLoS One. 2024. PMID: 39159214 Free PMC article.
-
COVID-19 and Mental Health: A "Pandemic Within a Pandemic".Adv Exp Med Biol. 2024;1458:1-18. doi: 10.1007/978-3-031-61943-4_1. Adv Exp Med Biol. 2024. PMID: 39102186 Review.
-
Loneliness, epigenetic age acceleration, and chronic health conditions.Psychol Aging. 2024 Jun;39(4):337-349. doi: 10.1037/pag0000822. Epub 2024 Apr 18. Psychol Aging. 2024. PMID: 38635160
-
The beneficial effects of social support and prosocial behavior on immunity and health: A psychoneuroimmunology perspective.Brain Behav Immun Health. 2024 Mar 18;37:100758. doi: 10.1016/j.bbih.2024.100758. eCollection 2024 May. Brain Behav Immun Health. 2024. PMID: 38524896 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
- P30-AG028748/AG/NIA NIH HHS/United States
- P51-OD011107/OD/NIH HHS/United States
- R01-AG034052/AG/NIA NIH HHS/United States
- P51 OD011107/OD/NIH HHS/United States
- P01-AG18911/AG/NIA NIH HHS/United States
- R01 DA024441/DA/NIDA NIH HHS/United States
- R01 AG043404/AG/NIA NIH HHS/United States
- R01-DA024441/DA/NIDA NIH HHS/United States
- P30 AG028748/AG/NIA NIH HHS/United States
- R37 AG033590/AG/NIA NIH HHS/United States
- P30 AG017265/AG/NIA NIH HHS/United States
- R01 AG034052/AG/NIA NIH HHS/United States
- P30-AG017265/AG/NIA NIH HHS/United States
- P01 AG018911/AG/NIA NIH HHS/United States
- R37-AG033590/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
