

GLP-1 stimulates insulin secretion by PKC-dependent TRPM4 and TRPM5 activation
- PMID: 26571400
- PMCID: PMC4665783
- DOI: 10.1172/JCI81975
GLP-1 stimulates insulin secretion by PKC-dependent TRPM4 and TRPM5 activation
Abstract
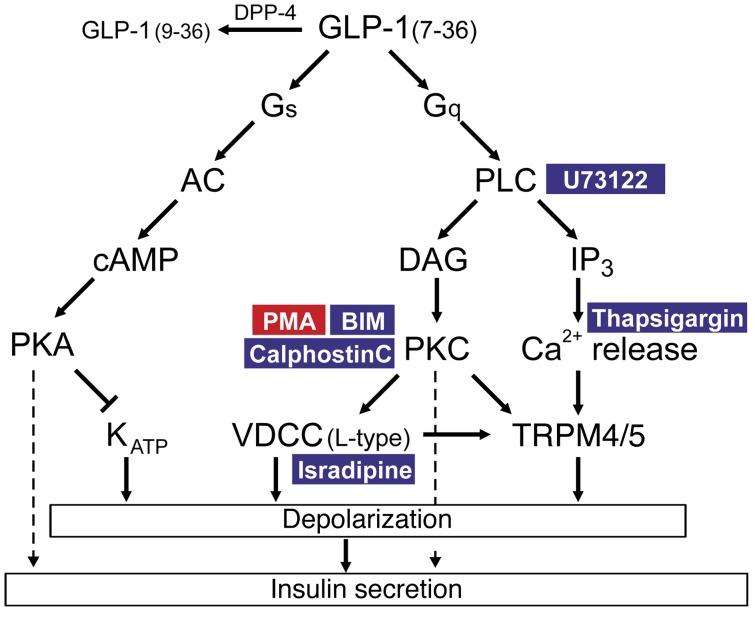
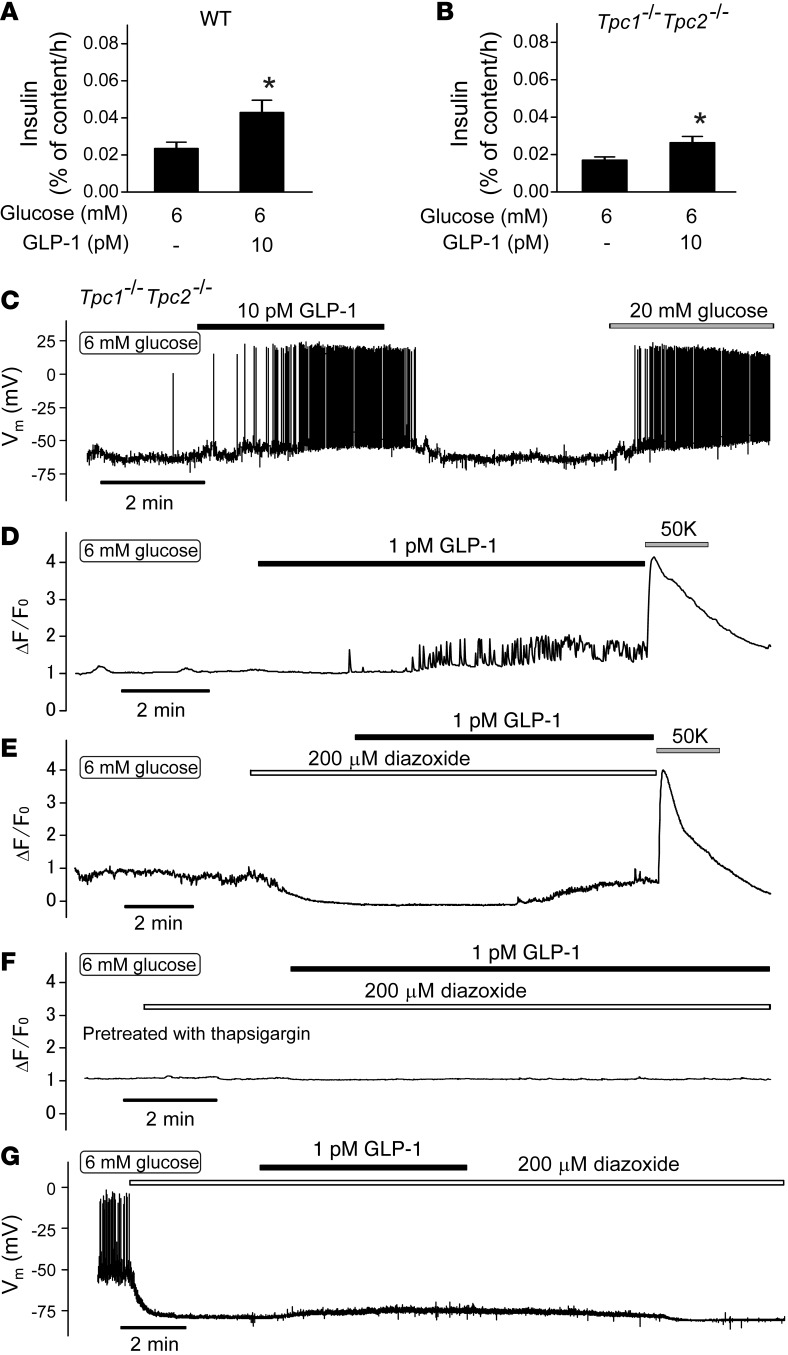
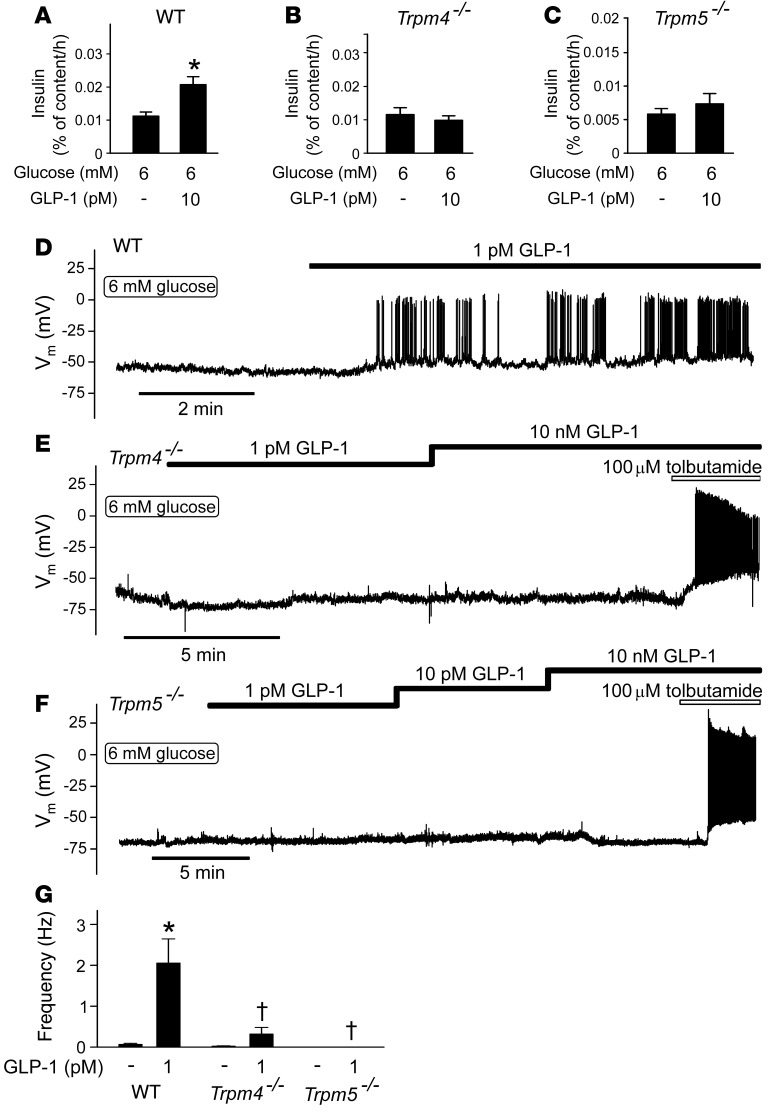
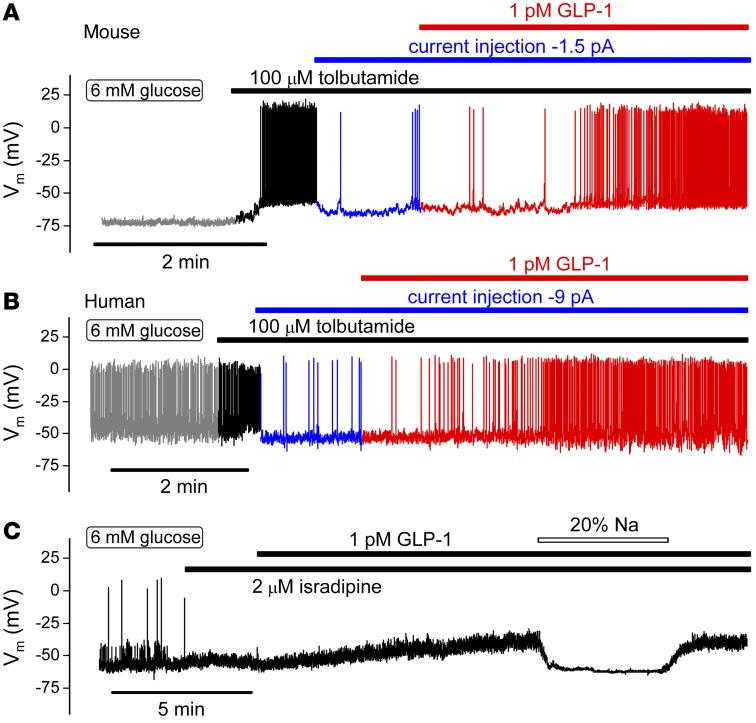
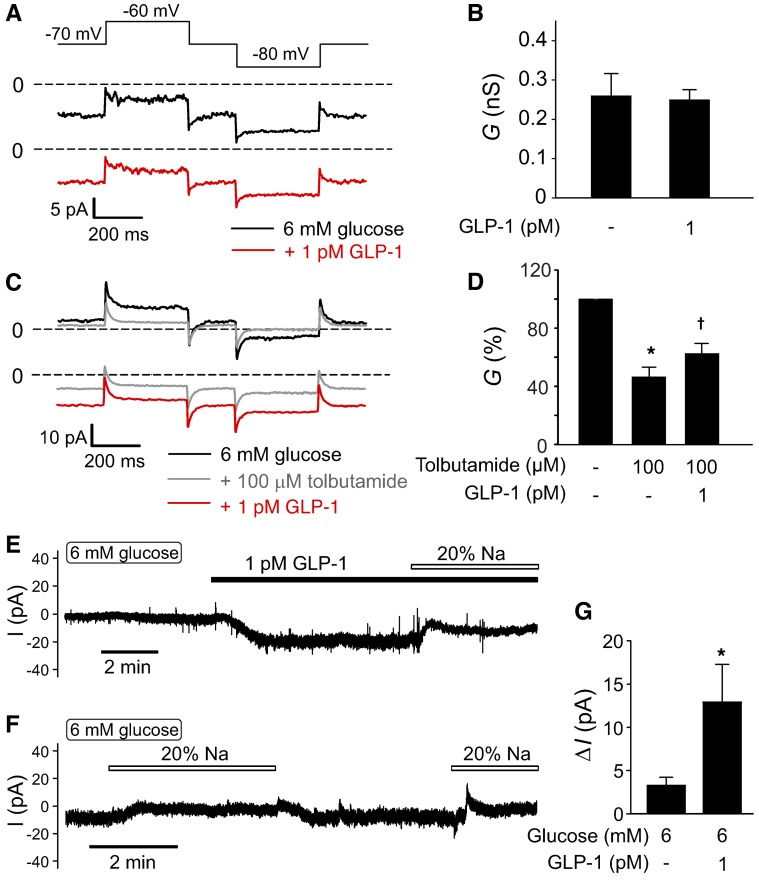
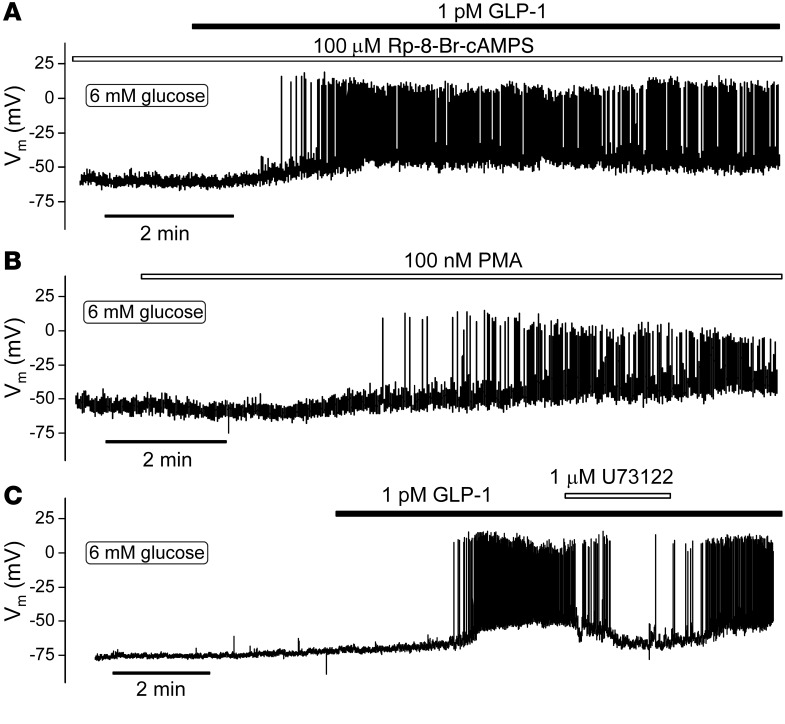
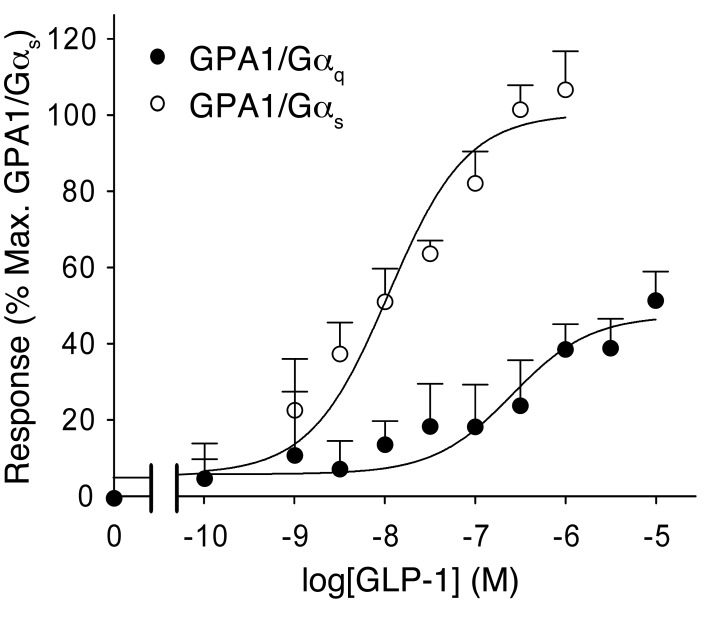
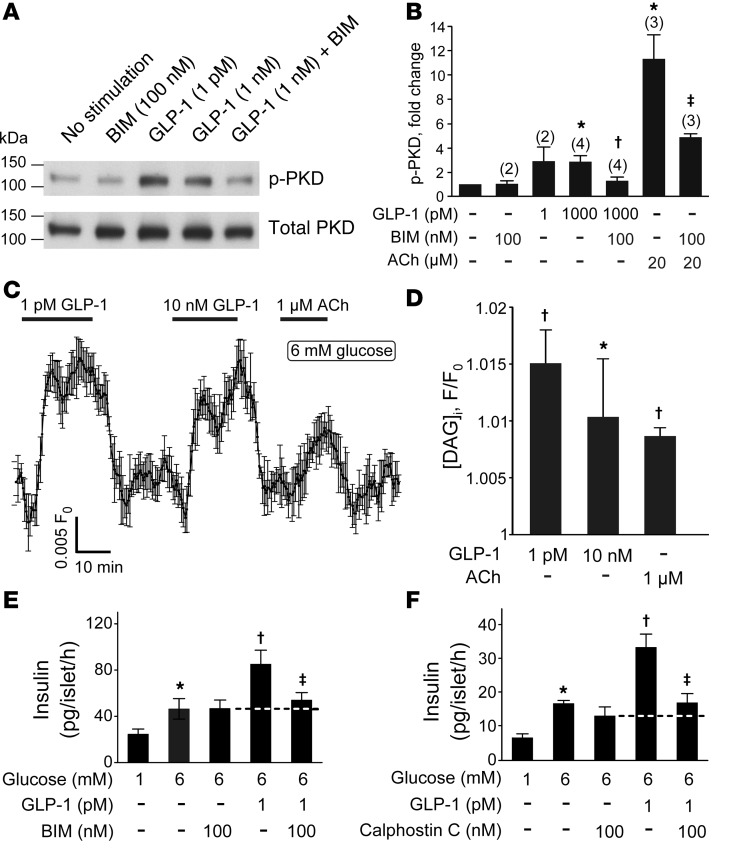
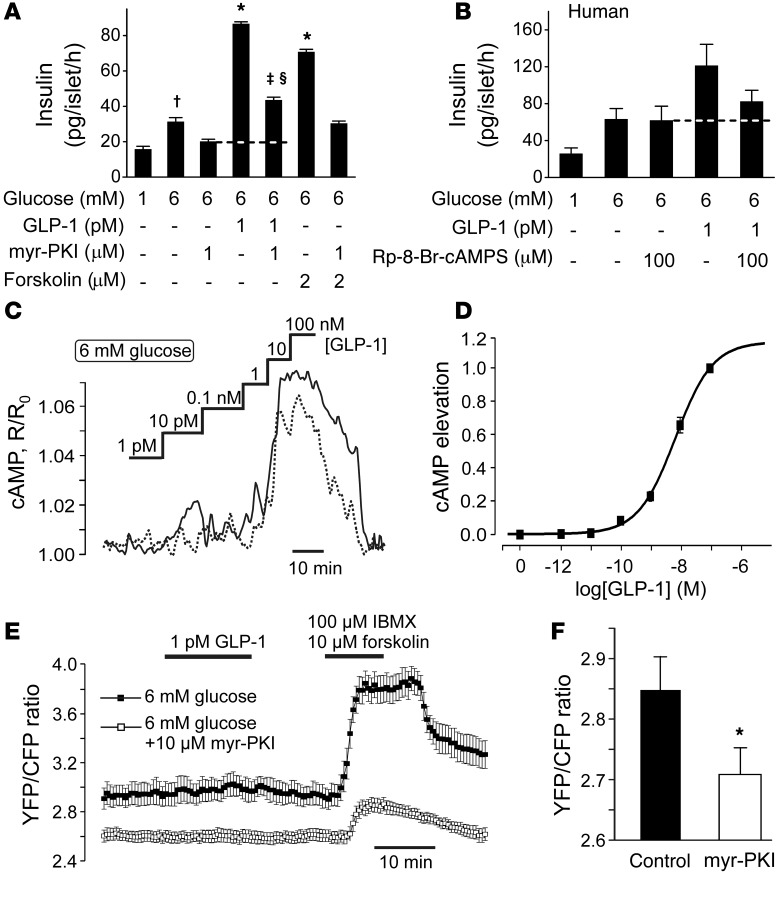
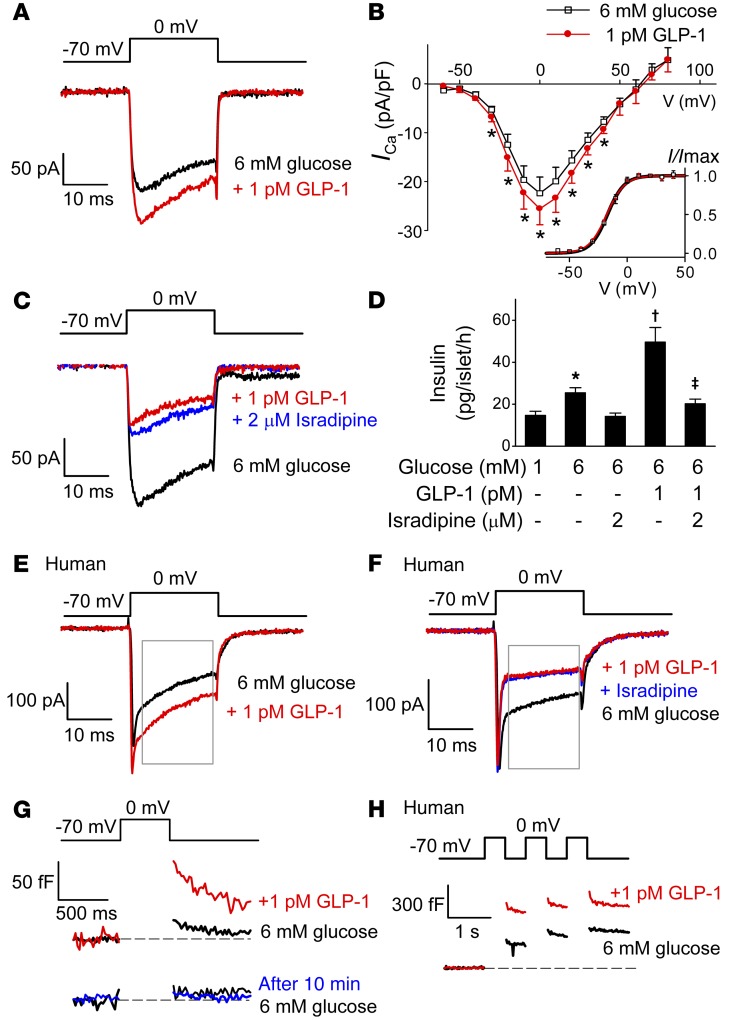
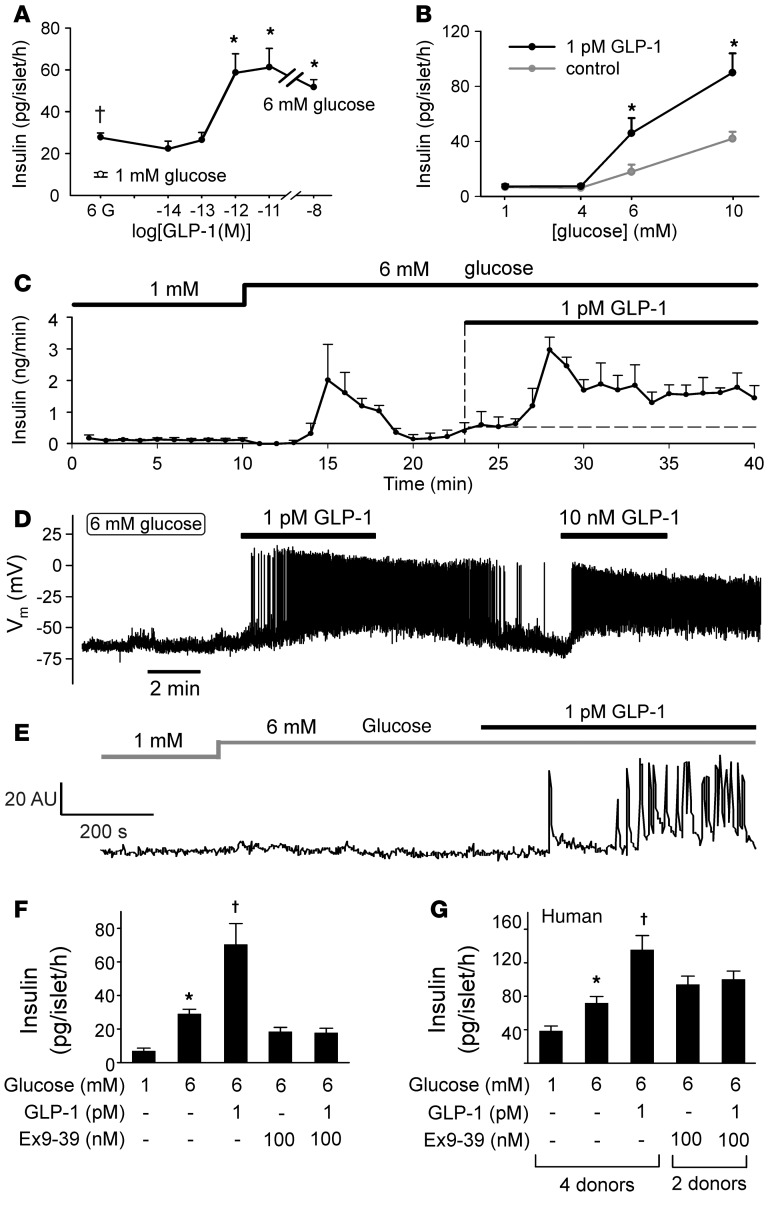
Strategies aimed at mimicking or enhancing the action of the incretin hormone glucagon-like peptide 1 (GLP-1) therapeutically improve glucose-stimulated insulin secretion (GSIS); however, it is not clear whether GLP-1 directly drives insulin secretion in pancreatic islets. Here, we examined the mechanisms by which GLP-1 stimulates insulin secretion in mouse and human islets. We found that GLP-1 enhances GSIS at a half-maximal effective concentration of 0.4 pM. Moreover, we determined that GLP-1 activates PLC, which increases submembrane diacylglycerol and thereby activates PKC, resulting in membrane depolarization and increased action potential firing and subsequent stimulation of insulin secretion. The depolarizing effect of GLP-1 on electrical activity was mimicked by the PKC activator PMA, occurred without activation of PKA, and persisted in the presence of PKA inhibitors, the KATP channel blocker tolbutamide, and the L-type Ca(2+) channel blocker isradipine; however, depolarization was abolished by lowering extracellular Na(+). The PKC-dependent effect of GLP-1 on membrane potential and electrical activity was mediated by activation of Na(+)-permeable TRPM4 and TRPM5 channels by mobilization of intracellular Ca(2+) from thapsigargin-sensitive Ca(2+) stores. Concordantly, GLP-1 effects were negligible in Trpm4 or Trpm5 KO islets. These data provide important insight into the therapeutic action of GLP-1 and suggest that circulating levels of this hormone directly stimulate insulin secretion by β cells.
Figures
Comment in
-
cAMP-independent effects of GLP-1 on β cells.J Clin Invest. 2015 Dec;125(12):4327-30. doi: 10.1172/JCI85004. Epub 2015 Nov 16. J Clin Invest. 2015. PMID: 26571393 Free PMC article.
Similar articles
-
A role of PLC/PKC-dependent pathway in GLP-1-stimulated insulin secretion.J Mol Med (Berl). 2017 Apr;95(4):361-368. doi: 10.1007/s00109-017-1508-6. Epub 2017 Jan 17. J Mol Med (Berl). 2017. PMID: 28097390 Review.
-
Cav1.2 and Cav1.3 are differentially coupled to glucagon-like peptide-1 potentiation of glucose-stimulated insulin secretion in the pancreatic beta-cell line INS-1.J Pharmacol Exp Ther. 2009 Nov;331(2):724-32. doi: 10.1124/jpet.109.158519. Epub 2009 Aug 26. J Pharmacol Exp Ther. 2009. PMID: 19710366 Free PMC article.
-
cAMP-independent effects of GLP-1 on β cells.J Clin Invest. 2015 Dec;125(12):4327-30. doi: 10.1172/JCI85004. Epub 2015 Nov 16. J Clin Invest. 2015. PMID: 26571393 Free PMC article.
-
Actions of glucagon-like peptide-1 on KATP channel-dependent and -independent effects of glucose, sulphonylureas and nateglinide.J Endocrinol. 2006 Sep;190(3):889-96. doi: 10.1677/joe.1.06949. J Endocrinol. 2006. PMID: 17003289
-
Molecular physiology of glucagon-like peptide-1 insulin secretagogue action in pancreatic β cells.Prog Biophys Mol Biol. 2011 Nov;107(2):236-47. doi: 10.1016/j.pbiomolbio.2011.07.005. Epub 2011 Jul 19. Prog Biophys Mol Biol. 2011. PMID: 21782840 Free PMC article. Review.
Cited by
-
Pharmacological and Clinical Studies of Medicinal Plants That Inhibit Dipeptidyl Peptidase-IV.Drug Des Devel Ther. 2023 Nov 23;17:3473-3491. doi: 10.2147/DDDT.S426870. eCollection 2023. Drug Des Devel Ther. 2023. PMID: 38024536 Free PMC article. Review.
-
Molecular Mechanism of Pancreatic β-Cell Failure in Type 2 Diabetes Mellitus.Biomedicines. 2022 Mar 31;10(4):818. doi: 10.3390/biomedicines10040818. Biomedicines. 2022. PMID: 35453568 Free PMC article. Review.
-
Glucagon-like peptide 1 (GLP-1).Mol Metab. 2019 Dec;30:72-130. doi: 10.1016/j.molmet.2019.09.010. Epub 2019 Sep 30. Mol Metab. 2019. PMID: 31767182 Free PMC article. Review.
-
Allelic variant in the glucagon-like peptide 1 receptor gene associated with greater effect of liraglutide and exenatide on gastric emptying: A pilot pharmacogenetics study.Neurogastroenterol Motil. 2018 Jul;30(7):e13313. doi: 10.1111/nmo.13313. Epub 2018 Feb 28. Neurogastroenterol Motil. 2018. PMID: 29488276 Free PMC article. Clinical Trial.
-
Ca2+ Regulation of TRP Ion Channels.Int J Mol Sci. 2018 Apr 23;19(4):1256. doi: 10.3390/ijms19041256. Int J Mol Sci. 2018. PMID: 29690581 Free PMC article. Review.
References
-
- Danaei G, et al. National, regional, and global trends in fasting plasma glucose and diabetes prevalence since 1980: systematic analysis of health examination surveys and epidemiological studies with 370 country-years and 2.7 million participants. Lancet. 2011;378(9785):31–40. doi: 10.1016/S0140-6736(11)60679-X. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
