

Myeloid Cell-Restricted Insulin/IGF-1 Receptor Deficiency Protects against Skin Inflammation
- PMID: 26519530
- PMCID: PMC4789495
- DOI: 10.4049/jimmunol.1501237
Myeloid Cell-Restricted Insulin/IGF-1 Receptor Deficiency Protects against Skin Inflammation
Abstract
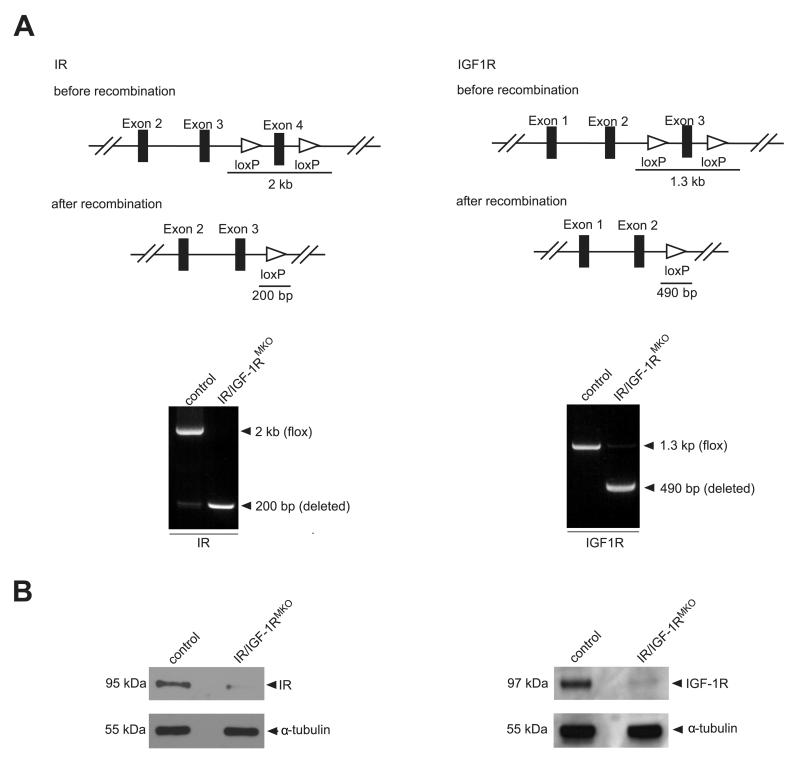
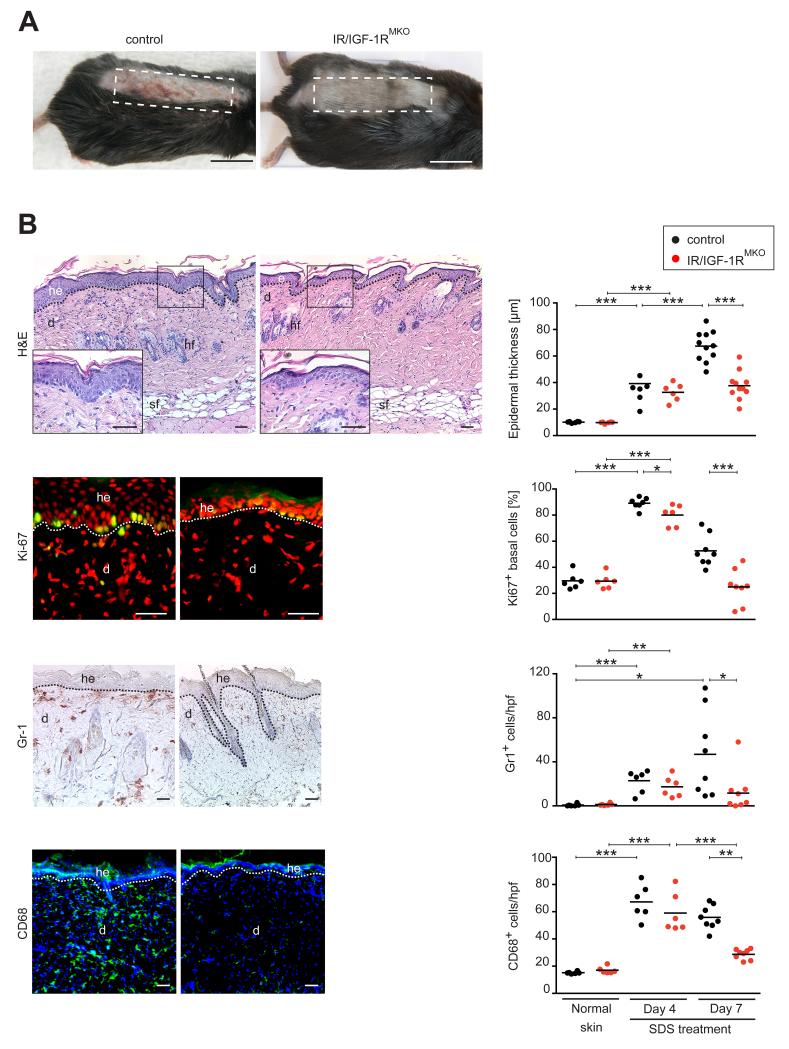
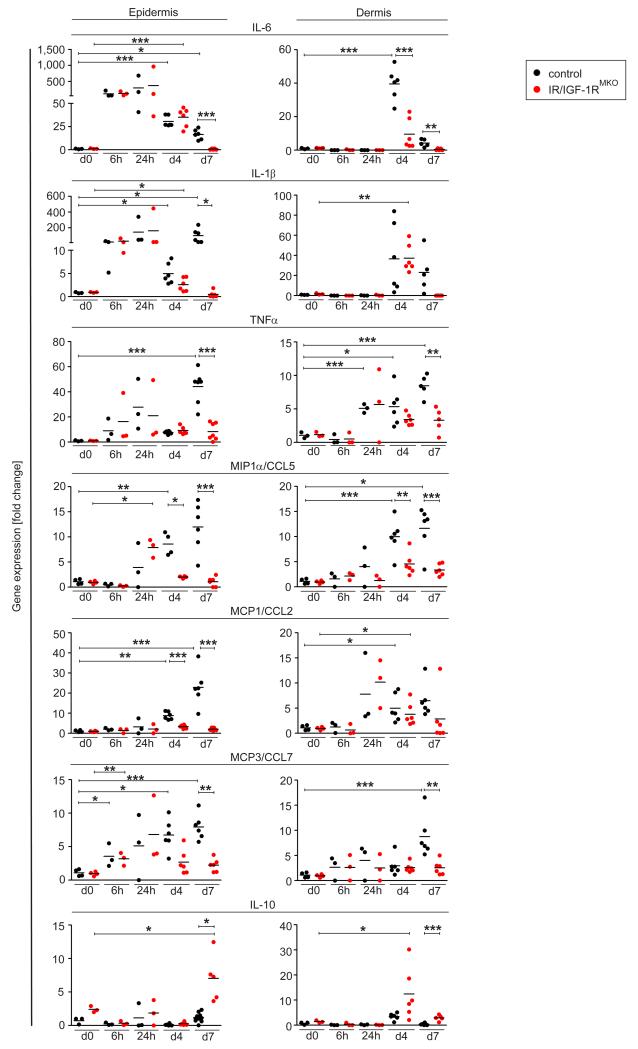
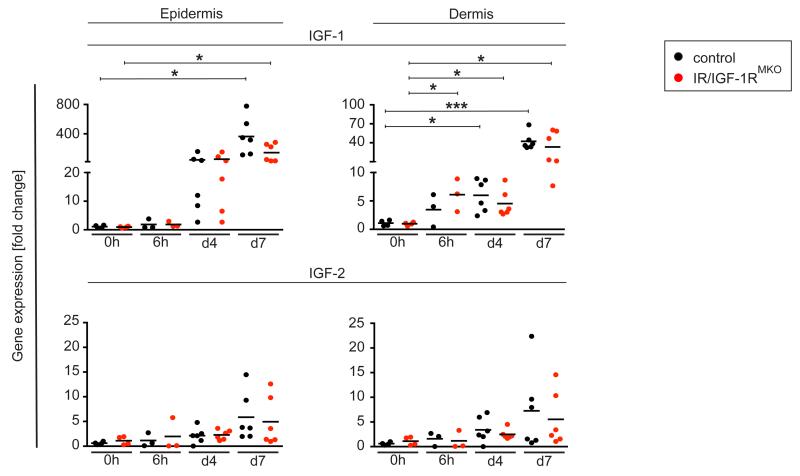
Myeloid cells are key regulators of tissue homeostasis and disease. Alterations in cell-autonomous insulin/IGF-1 signaling in myeloid cells have recently been implicated in the development of systemic inflammation and insulin-resistant diabetes mellitus type 2 (DM). Impaired wound healing and inflammatory skin diseases are frequent DM-associated skin pathologies, yet the underlying mechanisms are elusive. In this study, we investigated whether myeloid cell-restricted IR/IGF-1R signaling provides a pathophysiologic link between systemic insulin resistance and the development of cutaneous inflammation. Therefore, we generated mice lacking both the insulin and IGF-1 receptor in myeloid cells (IR/IGF-1R(MKO)). Whereas the kinetics of wound closure following acute skin injury was similar in control and IR/IGF-1R(MKO) mice, in two different conditions of dermatitis either induced by repetitive topical applications of the detergent SDS or by high-dose UV B radiation, IR/IGF-1R(MKO) mice were protected from inflammation, whereas controls developed severe skin dermatitis. Notably, whereas during the early phase in both inflammatory conditions the induction of epidermal proinflammatory cytokine expression was similar in control and IR/IGF-1R(MKO) mice, during the late stage, epidermal cytokine expression was sustained in controls but virtually abrogated in IR/IGF-1R(MKO) mice. This distinct kinetic of epidermal cytokine expression was paralleled by proinflammatory macrophage activation in controls and a noninflammatory phenotype in mutants. Collectively, our findings provide evidence for a proinflammatory IR/IGF-1R-dependent pathway in myeloid cells that plays a critical role in the dynamics of an epidermal-dermal cross-talk in cutaneous inflammatory responses, and may add to the mechanistic understanding of diseases associated with disturbances in myeloid cell IR/IGF-1R signaling, including DM.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures

Similar articles
-
Insulin-like growth factor-1 promotes wound healing in estrogen-deprived mice: new insights into cutaneous IGF-1R/ERα cross talk.J Invest Dermatol. 2012 Dec;132(12):2838-48. doi: 10.1038/jid.2012.228. Epub 2012 Jul 19. J Invest Dermatol. 2012. PMID: 22810305
-
The insulin-like growth factor-1 receptor is a negative regulator of nitric oxide bioavailability and insulin sensitivity in the endothelium.Diabetes. 2011 Aug;60(8):2169-78. doi: 10.2337/db11-0197. Epub 2011 Jun 15. Diabetes. 2011. PMID: 21677284 Free PMC article.
-
Thyrocyte-specific deletion of insulin and IGF-1 receptors induces papillary thyroid carcinoma-like lesions through EGFR pathway activation.Int J Cancer. 2018 Nov 15;143(10):2458-2469. doi: 10.1002/ijc.31779. Epub 2018 Sep 22. Int J Cancer. 2018. PMID: 30070361 Free PMC article.
-
Non-Coding RNAs in IGF-1R Signaling Regulation: The Underlying Pathophysiological Link between Diabetes and Cancer.Cells. 2019 Dec 14;8(12):1638. doi: 10.3390/cells8121638. Cells. 2019. PMID: 31847392 Free PMC article. Review.
-
Role of Interleukin-6 in Development of Insulin Resistance and Type 2 Diabetes Mellitus.Crit Rev Eukaryot Gene Expr. 2017;27(3):229-236. doi: 10.1615/CritRevEukaryotGeneExpr.2017019712. Crit Rev Eukaryot Gene Expr. 2017. PMID: 29199608 Review.
Cited by
-
Regulation of the immune system by the insulin receptor in health and disease.Front Endocrinol (Lausanne). 2023 Mar 13;14:1128622. doi: 10.3389/fendo.2023.1128622. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36992811 Free PMC article. Review.
-
Metabolic Regulation of Macrophage Activation.J Innate Immun. 2022;14(1):51-68. doi: 10.1159/000516780. Epub 2021 Jul 9. J Innate Immun. 2022. PMID: 34247159 Free PMC article. Review.
-
Isolation of macrophages from mouse skin wounds for single-cell RNA sequencing.STAR Protoc. 2022 Apr 19;3(2):101337. doi: 10.1016/j.xpro.2022.101337. eCollection 2022 Jun 17. STAR Protoc. 2022. PMID: 35496783 Free PMC article.
-
Macrophages play a key role in tissue repair and regeneration.PeerJ. 2022 Sep 29;10:e14053. doi: 10.7717/peerj.14053. eCollection 2022. PeerJ. 2022. PMID: 36196399 Free PMC article. Review.
-
Remodeling of whole-body lipid metabolism and a diabetic-like phenotype caused by loss of CDK1 and hepatocyte division.Elife. 2020 Dec 21;9:e63835. doi: 10.7554/eLife.63835. Elife. 2020. PMID: 33345777 Free PMC article.
References
-
- McNelis JC, Olefsky JM. Macrophages, Immunity, and Metabolic Disease. Immunity. 2014;1:36–48. - PubMed
-
- Baumgartl J, Baudler S, Scherner M, Babaev V, Makowski L, Suttles J, McDuffie M, Tobe K, Kadowaki T, Fazio S, Kahn CR, Hotamisligil GS, Krone W, Linton M, Brüning JC. Myeloid lineage cell-restricted insulin resistance protects apolipoproteinE-deficient mice against atherosclerosis. Cell. Metab. 2006;3:247–256. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous