

Convergence of Acquired Mutations and Alternative Splicing of CD19 Enables Resistance to CART-19 Immunotherapy
- PMID: 26516065
- PMCID: PMC4670800
- DOI: 10.1158/2159-8290.CD-15-1020
Convergence of Acquired Mutations and Alternative Splicing of CD19 Enables Resistance to CART-19 Immunotherapy
Abstract
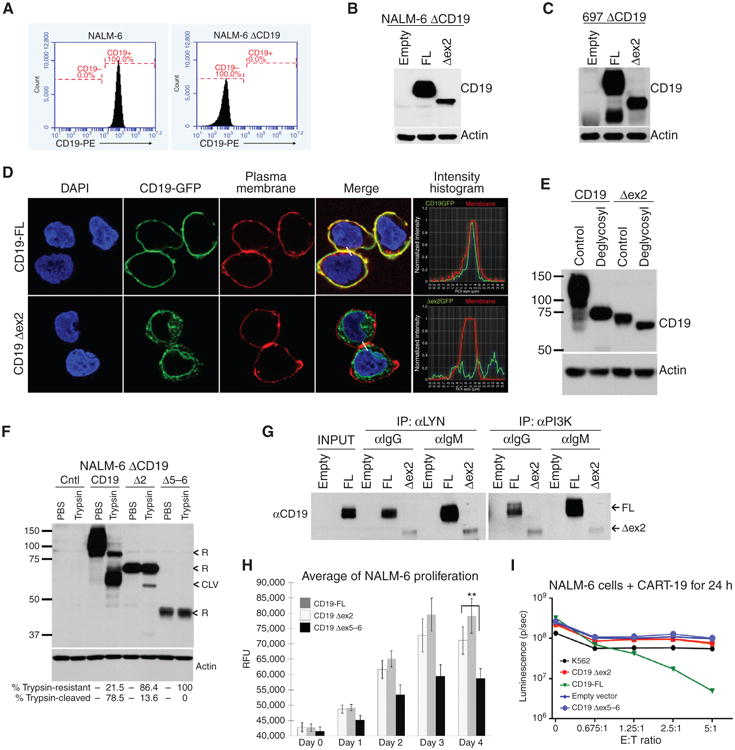
The CD19 antigen, expressed on most B-cell acute lymphoblastic leukemias (B-ALL), can be targeted with chimeric antigen receptor-armed T cells (CART-19), but relapses with epitope loss occur in 10% to 20% of pediatric responders. We detected hemizygous deletions spanning the CD19 locus and de novo frameshift and missense mutations in exon 2 of CD19 in some relapse samples. However, we also discovered alternatively spliced CD19 mRNA species, including one lacking exon 2. Pull-down/siRNA experiments identified SRSF3 as a splicing factor involved in exon 2 retention, and its levels were lower in relapsed B-ALL. Using genome editing, we demonstrated that exon 2 skipping bypasses exon 2 mutations in B-ALL cells and allows expression of the N-terminally truncated CD19 variant, which fails to trigger killing by CART-19 but partly rescues defects associated with CD19 loss. Thus, this mechanism of resistance is based on a combination of deleterious mutations and ensuing selection for alternatively spliced RNA isoforms.
Significance: CART-19 yield 70% response rates in patients with B-ALL, but also produce escape variants. We discovered that the underlying mechanism is the selection for preexisting alternatively spliced CD19 isoforms with the compromised CART-19 epitope. This mechanism suggests a possibility of targeting alternative CD19 ectodomains, which could improve survival of patients with B-cell neoplasms.
©2015 American Association for Cancer Research.
Conflict of interest statement
Figures
Comment in
-
Overcoming Antigen Escape with CAR T-cell Therapy.Cancer Discov. 2015 Dec;5(12):1238-40. doi: 10.1158/2159-8290.CD-15-1275. Cancer Discov. 2015. PMID: 26637657 Free PMC article.
Similar articles
-
CD19 Isoforms Enabling Resistance to CART-19 Immunotherapy Are Expressed in B-ALL Patients at Initial Diagnosis.J Immunother. 2017 Jun;40(5):187-195. doi: 10.1097/CJI.0000000000000169. J Immunother. 2017. PMID: 28441264 Free PMC article.
-
High-throughput mutagenesis identifies mutations and RNA-binding proteins controlling CD19 splicing and CART-19 therapy resistance.Nat Commun. 2022 Sep 22;13(1):5570. doi: 10.1038/s41467-022-31818-y. Nat Commun. 2022. PMID: 36138008 Free PMC article.
-
Genetic Mechanism of Leukemia Relapse Following CD19 Chimeric Antigen Receptor T Cell Therapy.Cancer Biother Radiopharm. 2022 Jun;37(5):335-341. doi: 10.1089/cbr.2020.4630. Epub 2021 Mar 18. Cancer Biother Radiopharm. 2022. PMID: 33739864 Review.
-
Chimeric Antigen Receptor Therapy in Acute Lymphoblastic Leukemia Clinical Practice.Curr Hematol Malig Rep. 2017 Aug;12(4):370-379. doi: 10.1007/s11899-017-0394-x. Curr Hematol Malig Rep. 2017. PMID: 28656487 Review.
-
Unraveling resistance mechanisms in anti-CD19 chimeric antigen receptor-T therapy for B-ALL: a novel in vitro model and insights into target antigen dynamics.J Transl Med. 2024 May 21;22(1):482. doi: 10.1186/s12967-024-05254-z. J Transl Med. 2024. PMID: 38773607 Free PMC article.
Cited by
-
Efficient combinatorial adaptor-mediated targeting of acute myeloid leukemia with CAR T-cells.Leukemia. 2024 Dec;38(12):2598-2613. doi: 10.1038/s41375-024-02409-1. Epub 2024 Sep 18. Leukemia. 2024. PMID: 39294295 Free PMC article.
-
High-content molecular profiling of T-cell therapy in oncology.Mol Ther Oncolytics. 2016 Mar 30;3:16009. doi: 10.1038/mto.2016.9. eCollection 2016. Mol Ther Oncolytics. 2016. PMID: 27626060 Free PMC article. Review.
-
Insights gained from single-cell analysis of chimeric antigen receptor T-cell immunotherapy in cancer.Mil Med Res. 2023 Nov 8;10(1):52. doi: 10.1186/s40779-023-00486-4. Mil Med Res. 2023. PMID: 37941075 Free PMC article. Review.
-
Overcoming Resistance to Immune Checkpoint Inhibitors in Head and Neck Squamous Cell Carcinomas.Front Oncol. 2021 Mar 5;11:596290. doi: 10.3389/fonc.2021.596290. eCollection 2021. Front Oncol. 2021. PMID: 33747915 Free PMC article. Review.
-
Dual CD19 and CD123 targeting prevents antigen-loss relapses after CD19-directed immunotherapies.J Clin Invest. 2016 Oct 3;126(10):3814-3826. doi: 10.1172/JCI87366. Epub 2016 Aug 29. J Clin Invest. 2016. PMID: 27571406 Free PMC article.
References
-
- Roberts KG, Mullighan CG. Genomics in acute lymphoblastic leukaemia: insights and treatment implications. Nat Rev Clin Oncol. 2015;12:344–57. - PubMed
-
- Topp MS, Gokbuget N, Zugmaier G, Klappers P, Stelljes M, Neumann S, et al. Phase II trial of the anti-CD19 bispecific T cell-engager blinatumomab shows hematologic and molecular remissions in patients with relapsed or refractory B-precursor acute lymphoblastic leukemia. J Clin Oncol. 2014;32:4134–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
