

Review
doi: 10.1101/cshperspect.a019356.
Long-Range Chromatin Interactions
Affiliations
- PMID: 26430217
- PMCID: PMC4588061
- DOI: 10.1101/cshperspect.a019356
Item in Clipboard
Review
Long-Range Chromatin Interactions
Cold Spring Harb Perspect Biol.
.
Abstract
To accommodate genomes in the limited space of the cell nucleus and ensure the correct execution of gene expression programs, genomes are packaged in complex fashion in the three-dimensional cell nucleus. As a consequence of the extensive higher-order organization of chromosomes, distantly located genomic regions on the same or distinct chromosomes undergo long-range interactions. This article discusses the nature of long interactions, mechanisms of their formation, and their emerging functional roles in gene regulation and genome maintenance.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

Chromosome territories and genes in three-dimensional (3D) space. Fluorescence in situ hybridization (FISH) visualizes the spatial organization of the genome. (A) Chromosomes exist in the form of chromosome territories in the interphase nucleus. The DNA of each chromosome occupies a spatially well-defined fraction of the nuclear volume, typically about 1–2 μm in diameter. Chromosome 11 (green) in the nucleus (blue) of MCF10A breast cancer cells is shown. (B) Individual genes appear as distinct spots. MYC (red) and TGFBR2 (green) in MCF10A cells are shown. (C) The position of a chromosome or a gene can be expressed as the distance from the center of the nucleus or relative to other genes. The distribution of chromosomes and genes is nonrandom with some chromosomes (red) preferentially occupying internal positions and others (green) occupying peripheral positions. The nonrandom radial positioning also gives rise to nonrandom genome neighborhoods. (D) The distribution of a gene is probabilistic. Mapping of the position of the IGH gene in human lymphocytes in several hundred individual cells shows that its distribution is distinct from a random distribution; however, the IGH locus can be found in variable locations in individual cells. Each red dot represents the position of an IGH allele in a single cell. (C, Modified from Meaburn and Misteli 2007.)
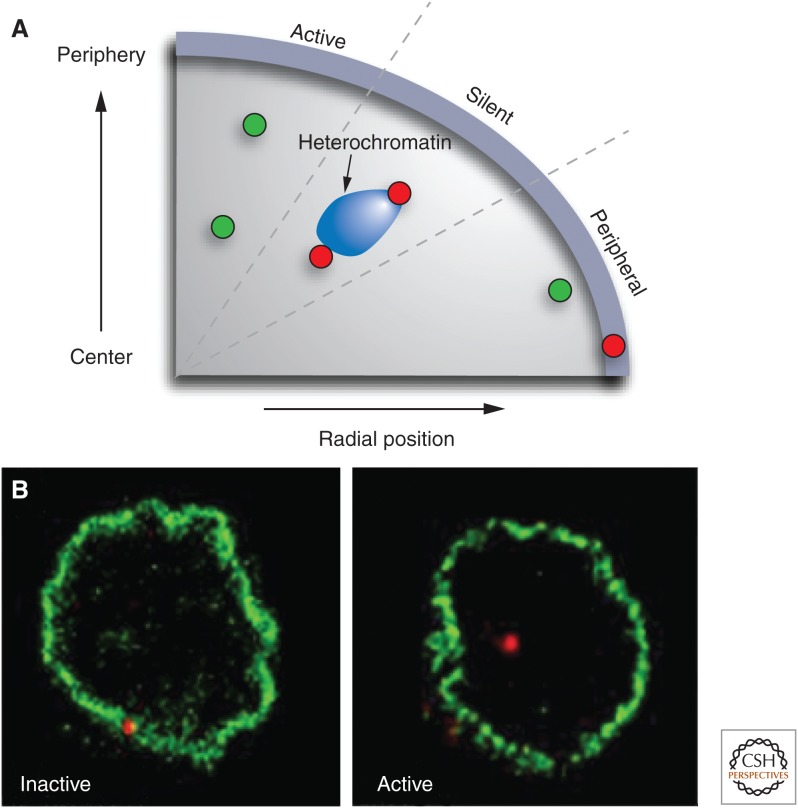
The role of the nuclear periphery in gene regulation. (A) Active genes (green) show a large range of radial positions; the precise radial position of a locus does not correlate with its activity level. Inactive genes (red) may associate with heterochromatin blocks at various radial positions. In contrast, physical association with the nuclear periphery is often linked to silencing. Genes that are in close proximity to the nuclear envelope but do not physically interact with it may be active. (B) Association of a gene locus with the nuclear edge often correlates with activity. The IgH locus (red) associates with the nuclear lamina when it is inactive in hematopoietic progenitor cells, but dissociates on activation in activated pro B cells. (A, Redrawn from Takizawa et al. 2008; B, reproduced, with permission, from Kosak et al. 2002, © AAAS.)

A view into the nuclear interior. Genomes exist in vivo in the form of chromosome territories made up of chromatin folded into still poorly understood higher-order fibers (multicolored areas). Chromatin fibers occupy the entire nuclear space, yet despite the presence of up to 2 m of DNA in a mammalian nucleus, there is considerable open nucleoplasmic space. Active genes (red) may loop out from chromosome territories and aggregate in centers of transcription (shaded areas), which contain appropriate transcription factors (blue, green). It has been suggested that coregulated genes cluster in shared transcription sites. (Reproduced from Misteli 2011; illustration by Anatomy Blue.)
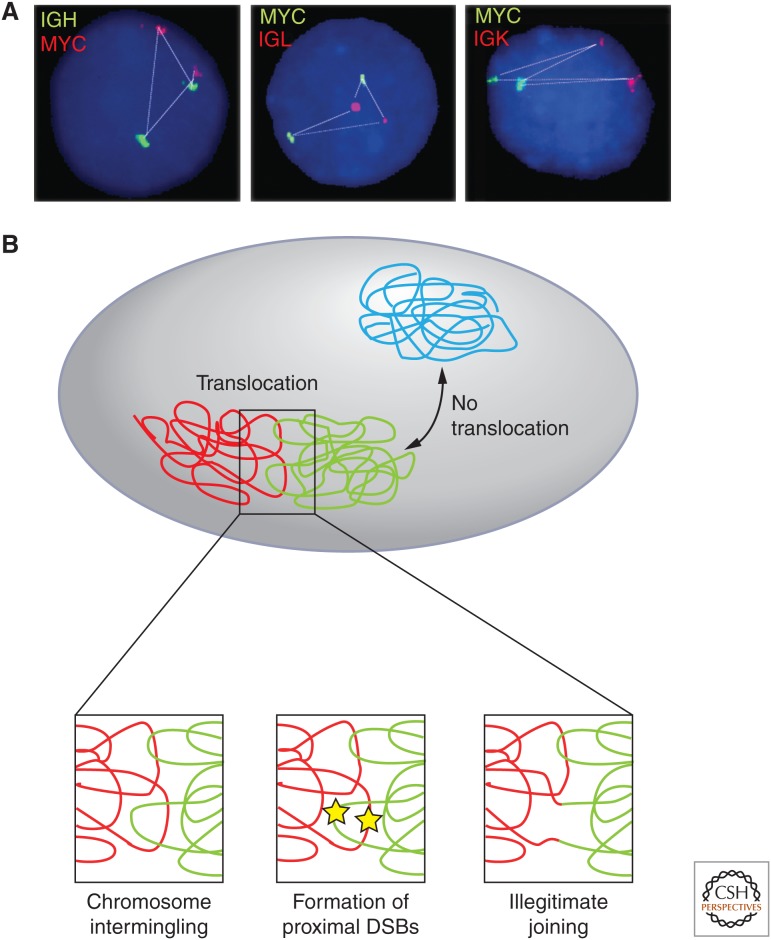
Roles of genome organization in determining chromosome translocations. The nonrandom organization of genes and chromosomes contributes to the formation of cancer translocations. (A) The physical distance of MYC to its translocation partners IGH, IGL, and IGK correlates with their translocations frequency (MYC-IGH > MYC-IGL > MYC-IGK). (B) Translocations preferentially occur between proximally positioned chromosomes (red, green), and only rarely between distally located chromosomes (blue). Closely juxtaposed double-strand breaks (yellow stars) occurring at the interface between chromosomes create free chromosome ends that may recombine to form a chromosome translocation by illegitimate joining. (A, Adapted from Roix et al. 2003; B, reproduced, with permission, from Misteli 2010, © Cold Spring Harbor Laboratory Press.)
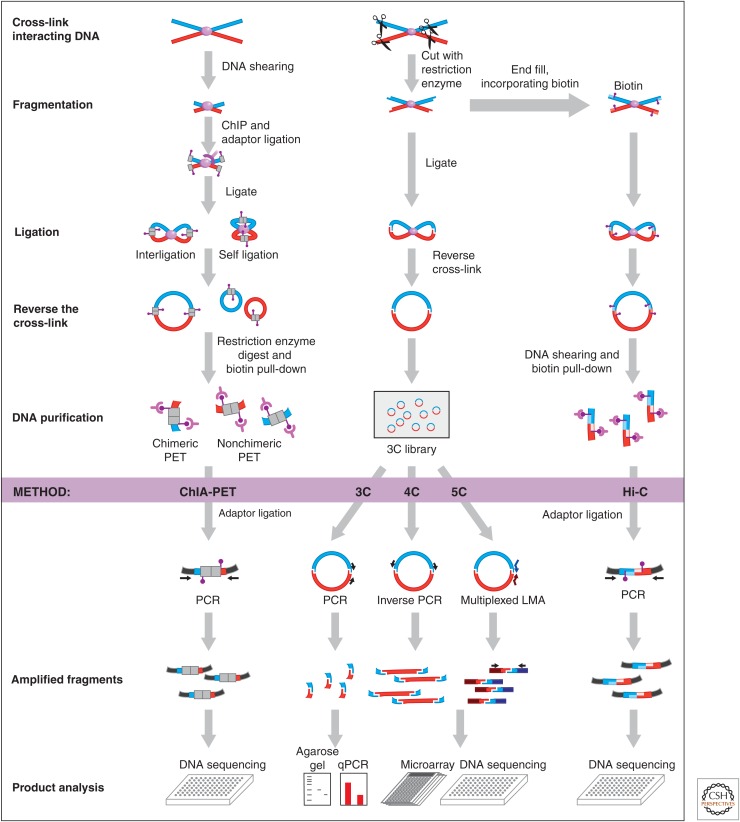
Overview of 3C-based methods. All 3C-based methods rely on covalently linking spatially proximal chromatin segments. Through a series of steps chromatin is then fragmented and religated and ligation products are detected using PCR, microarrays, or deep sequencing. In ChIA-PET (chromatin interaction analysis with paired-end tag sequencing), chromatin fragmentation is achieved by shearing and ligation junctions are marked by adaptors that contain recognition sites for type I restriction enzymes. Redigestion of ligation products with such enzymes yields small ligation, junction-containing molecules that can be analyzed by deep sequencing. 3C, 4C, and 5C use restriction enzyme digestion to fragment cross-linked chromatin. Religation of DNA then produces ligation products that can be directly analyzed by PCR (3C), inverse PCR (4C), or ligation-mediated amplification (LMA; 5C). Hi-C is as 3C, but includes a step to incorporate biotinylated nucleotides before religation. This facilitates purification of ligation junctions that are then analyzed by deep sequencing. (Adapted from Sanyal et al. 2011.)
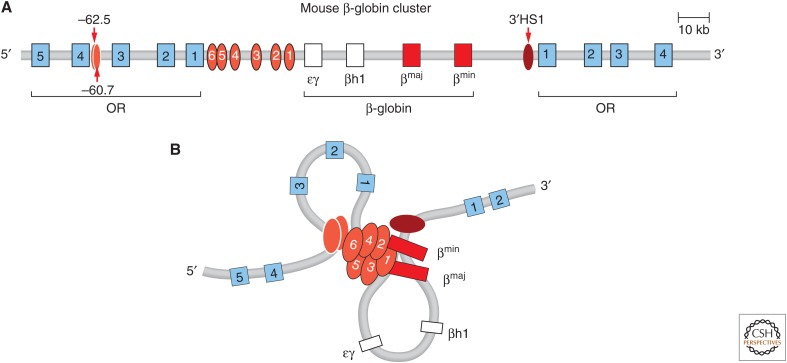
The mouse β-globin locus. (A) Schematic representation of the β-globin locus. (B) Looping between the LCR, the globin genes, and up- and downstream elements bound by CTCF (HS-62.5/-60.7 and 3′HS1) is observed in globin-expressing erythroid cells. (Redrawn from de Laat and Grosveld 2003, with permission from Springer Science and Business Media.)
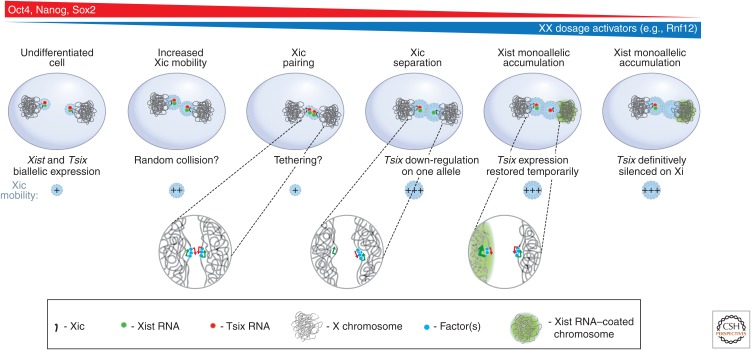
Interchromosomal interactions between Xics during early embryonic development. During early stages of embryonic stem (ES) cell differentiation, the two X chromosomes are highly mobile, possibly to allow pairing of the two Xics. During this pairing stage, through a process still largely unknown, one of the two X chromosomes will down-regulate expression of Tsix, a negative regulator of the Xist RNA gene. On that chromosome, Xist will be expressed and Xist RNA will accumulate resulting in X-inactivation. The other X chromosome will continue to express Tsix and the chromosome will remain active. (Redrawn, with permission, from Masui et al. 2011, © Elsevier.)
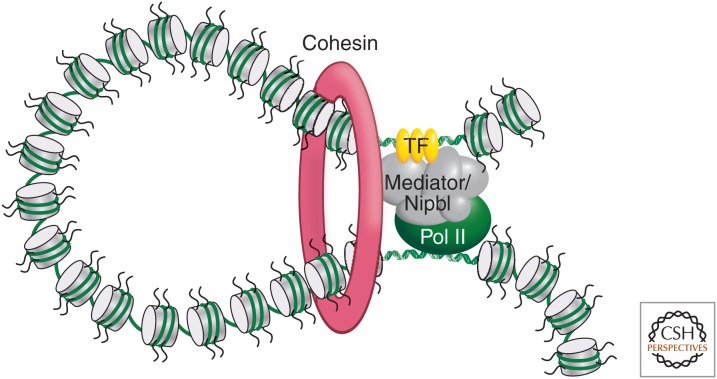
Cohesin and Mediator complexes mediate long-range interactions between promoters and enhancers. The Cohesin and Mediator have been shown to act together to form and stabilize chromatin looping interactions between gene promoters and distal enhancers (Kagey et al. 2010). A model is shown of how these complexes might mediate chromatin looping, although molecular details are currently unknown. (Redrawn, with permission, from Young 2011, © Elsevier.)
Similar articles
-
The contribution of nuclear compartmentalization to gene regulation.Cell. 2002 Feb 22;108(4):513-21. doi: 10.1016/s0092-8674(02)00650-5. Cell. 2002. PMID: 11909522 Review.
-
The cell nucleus taking centre stage. Workshop on the functional organization of the cell nucleus.EMBO Rep. 2006 Dec;7(12):1211-5. doi: 10.1038/sj.embor.7400840. Epub 2006 Oct 27. EMBO Rep. 2006. PMID: 17068488 Free PMC article. No abstract available.
-
Chromatin organization in the mammalian nucleus.Int Rev Cytol. 2005;242:283-336. doi: 10.1016/S0074-7696(04)42007-5. Int Rev Cytol. 2005. PMID: 15598472 Review.
-
The budding yeast nucleus.Cold Spring Harb Perspect Biol. 2010 Aug;2(8):a000612. doi: 10.1101/cshperspect.a000612. Epub 2010 Jun 16. Cold Spring Harb Perspect Biol. 2010. PMID: 20554704 Free PMC article. Review.
-
Functional architecture in the cell nucleus.Biochem J. 2001 Jun 1;356(Pt 2):297-310. doi: 10.1042/0264-6021:3560297. Biochem J. 2001. PMID: 11368755 Free PMC article. Review.
Cited by
-
Cohesin prevents cross-domain gene coactivation.Nat Genet. 2024 Aug;56(8):1654-1664. doi: 10.1038/s41588-024-01852-1. Epub 2024 Jul 24. Nat Genet. 2024. PMID: 39048795 Free PMC article.
-
The Self-Organizing Genome: Principles of Genome Architecture and Function.Cell. 2020 Oct 1;183(1):28-45. doi: 10.1016/j.cell.2020.09.014. Epub 2020 Sep 24. Cell. 2020. PMID: 32976797 Free PMC article. Review.
-
LDB1 establishes multi-enhancer networks to regulate gene expression.bioRxiv [Preprint]. 2024 Aug 24:2024.08.23.609430. doi: 10.1101/2024.08.23.609430. bioRxiv. 2024. Update in: Mol Cell. 2025 Jan 16;85(2):376-393.e9. doi: 10.1016/j.molcel.2024.11.037. PMID: 39229045 Free PMC article. Updated. Preprint.
-
Regulating Methylation at H3K27: A Trick or Treat for Cancer Cell Plasticity.Cancers (Basel). 2020 Sep 29;12(10):2792. doi: 10.3390/cancers12102792. Cancers (Basel). 2020. PMID: 33003334 Free PMC article. Review.
-
Programmed DNA elimination in the parasitic nematode Ascaris.PLoS Pathog. 2023 Feb 2;19(2):e1011087. doi: 10.1371/journal.ppat.1011087. eCollection 2023 Feb. PLoS Pathog. 2023. PMID: 36730159 Free PMC article. Review.
References
-
- Augui S, Filion GJ, Huart S, Nora E, Guggiari M, Maresca M, Stewart AF, Heard E. 2007. Sensing X chromosome pairs before X inactivation via a novel X-pairing region of the Xic. Science 318: 1632–1636. - PubMed
-
- Bacher CP, Guggiari M, Brors B, Augui S, Clerc P, Avner P, Eils R, Heard E. 2006. Transient colocalization of X-inactivation centres accompanies the initiation of X inactivation. Nat Cell Biol 8: 293–299. - PubMed
-
- Bantignies F, Roure V, Comet I, Leblanc B, Schuettengruber B, Bonnet J, Tixier V, Mas A, Cavalli G. 2011. Polycomb-dependent regulatory contacts between distant Hox loci in Drosophila. Cell 144: 214–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials